

MicroRNAs Induce a Permissive Chromatin Environment that Enables Neuronal Subtype-Specific Reprogramming of Adult Human Fibroblasts
- PMID: 28886366
- PMCID: PMC5679239
- DOI: 10.1016/j.stem.2017.08.002
MicroRNAs Induce a Permissive Chromatin Environment that Enables Neuronal Subtype-Specific Reprogramming of Adult Human Fibroblasts
Abstract
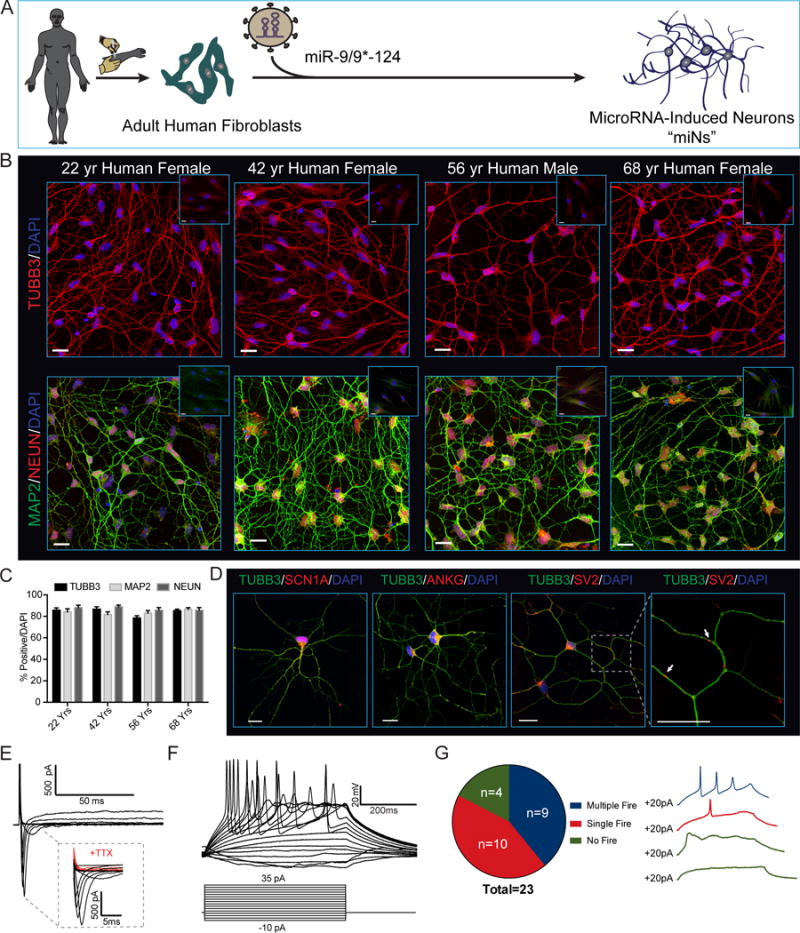
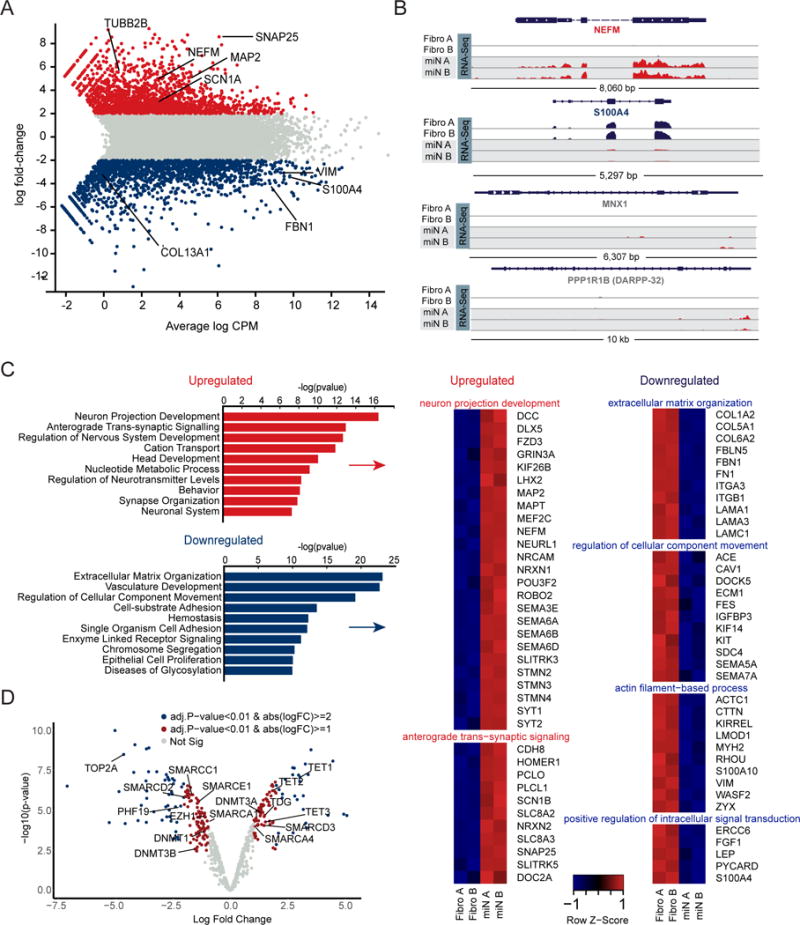
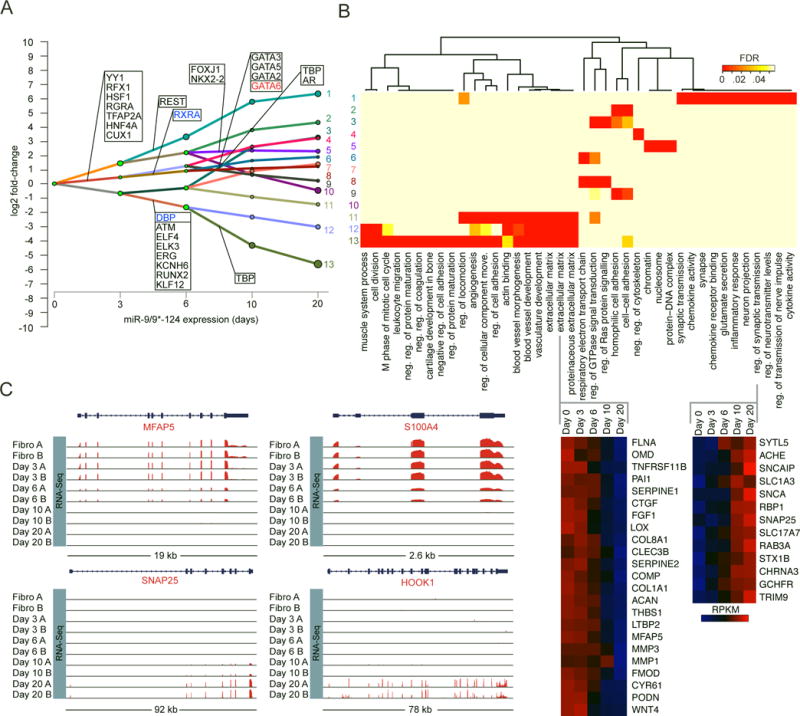
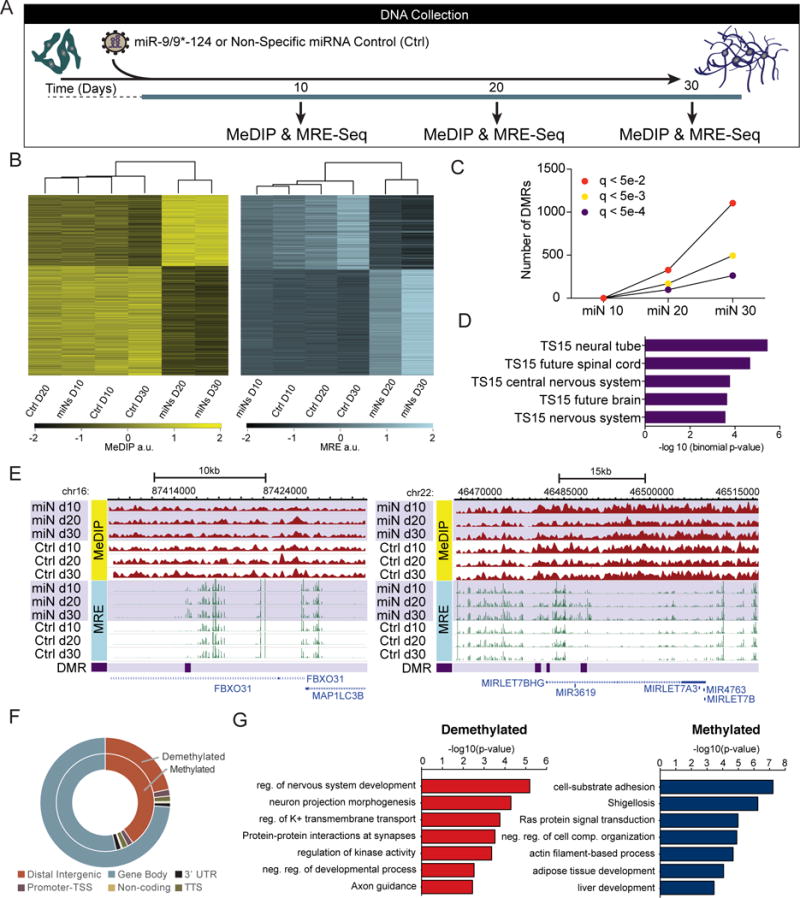
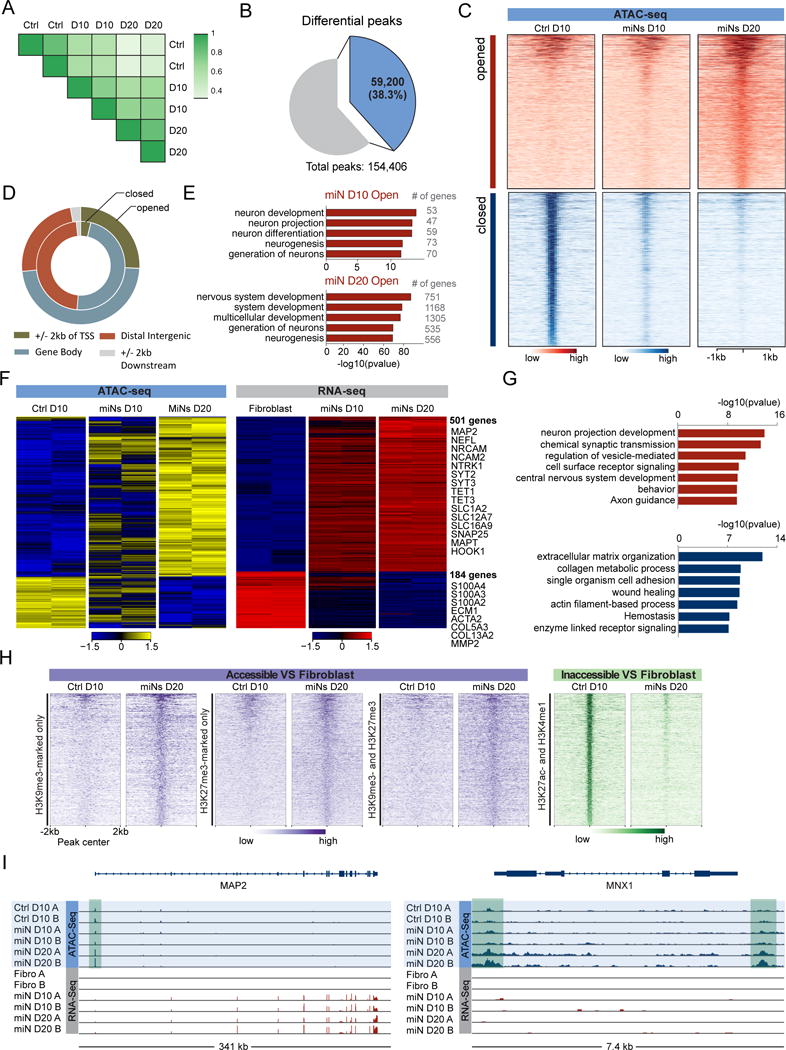
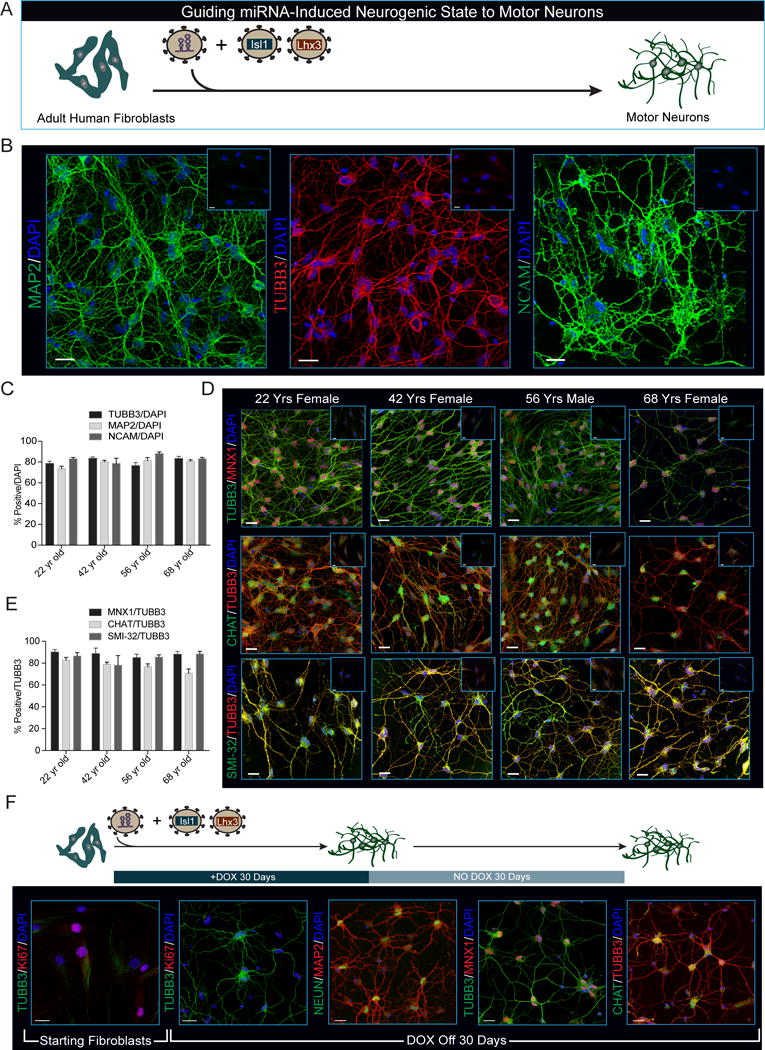
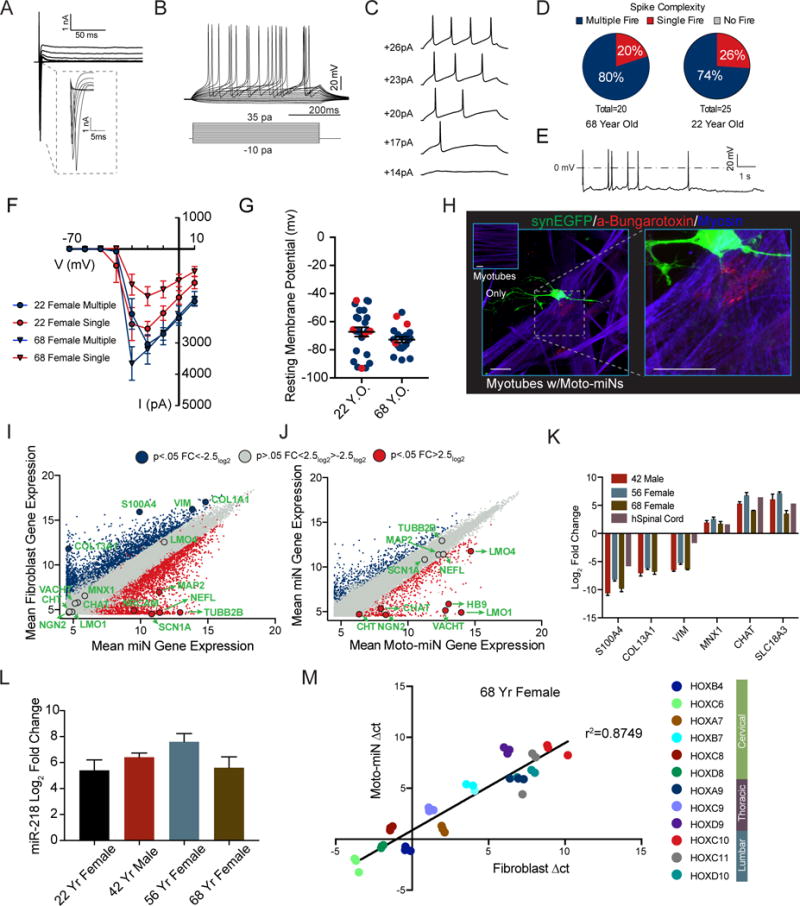
Directed reprogramming of human fibroblasts into fully differentiated neurons requires massive changes in epigenetic and transcriptional states. Induction of a chromatin environment permissive for acquiring neuronal subtype identity is therefore a major barrier to fate conversion. Here we show that the brain-enriched miRNAs miR-9/9∗ and miR-124 (miR-9/9∗-124) trigger reconfiguration of chromatin accessibility, DNA methylation, and mRNA expression to induce a default neuronal state. miR-9/9∗-124-induced neurons (miNs) are functionally excitable and uncommitted toward specific subtypes but possess open chromatin at neuronal subtype-specific loci, suggesting that such identity can be imparted by additional lineage-specific transcription factors. Consistently, we show that ISL1 and LHX3 selectively drive conversion to a highly homogeneous population of human spinal cord motor neurons. This study shows that modular synergism between miRNAs and neuronal subtype-specific transcription factors can drive lineage-specific neuronal reprogramming, providing a general platform for high-efficiency generation of distinct subtypes of human neurons.
Keywords: DNA methylation; cell fate; chromatin accessibility; chromatin remodeling; direct reprogramming; epigenetics; human neurons; microRNA; motor neurons; neurogenesis.
Copyright © 2017 Elsevier Inc. All rights reserved.
Conflict of interest statement
These authors contributed equally to this work
Figures
References
-
- Cantone I, Fisher AG. Epigenetic programming and reprogramming during development. Nat Struct Mol Biol. 2013;20:282–289. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
