

Silencing of retrotransposons by SETDB1 inhibits the interferon response in acute myeloid leukemia
- PMID: 28887438
- PMCID: PMC5674883
- DOI: 10.1083/jcb.201612160
Silencing of retrotransposons by SETDB1 inhibits the interferon response in acute myeloid leukemia
Abstract
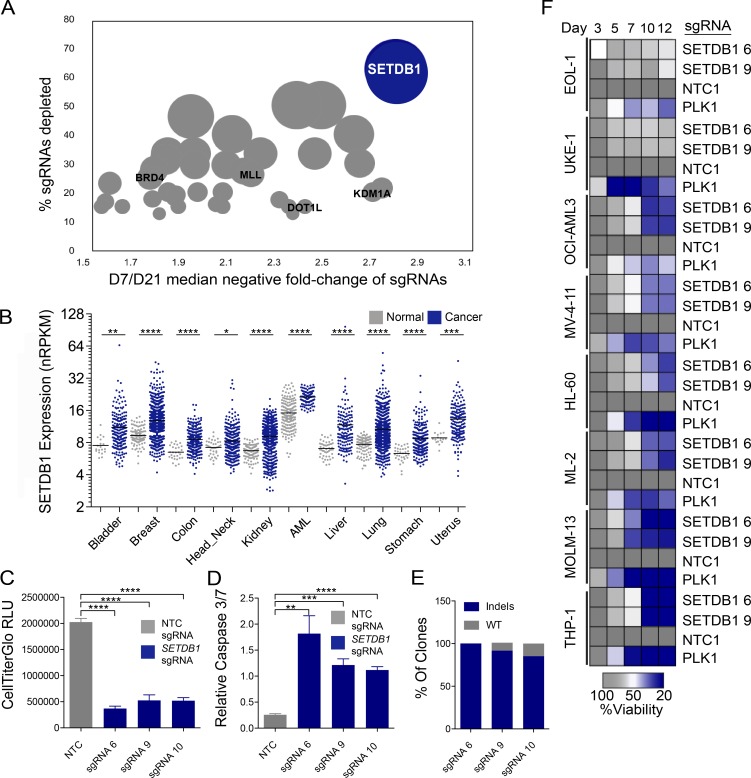
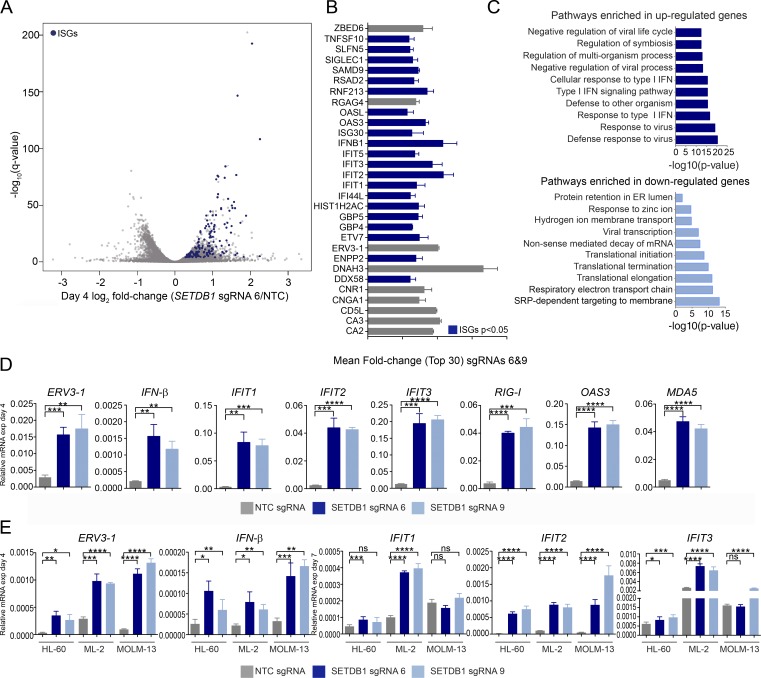
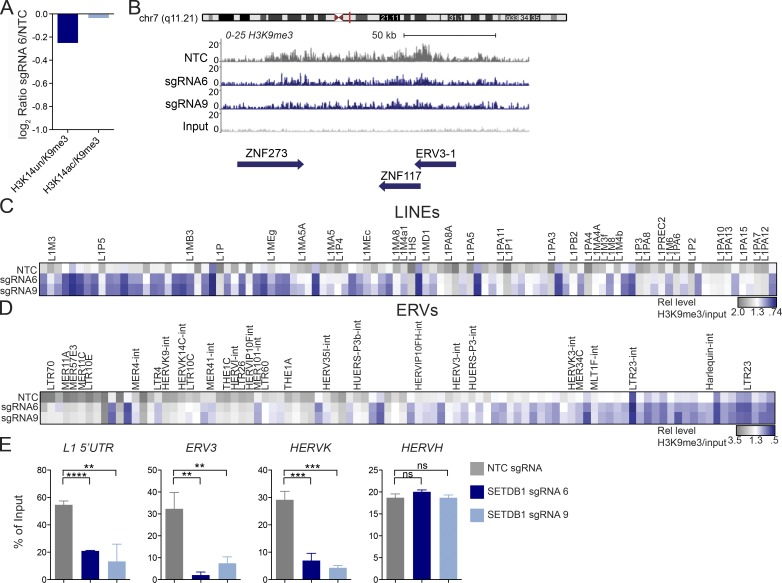
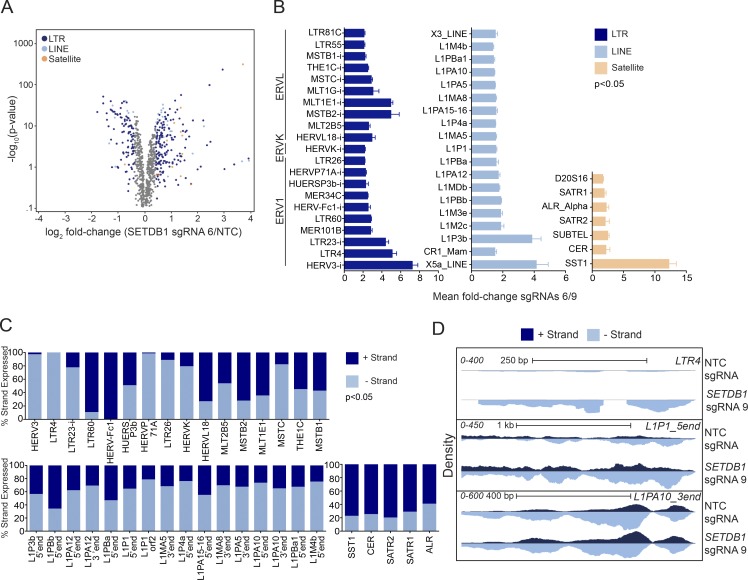
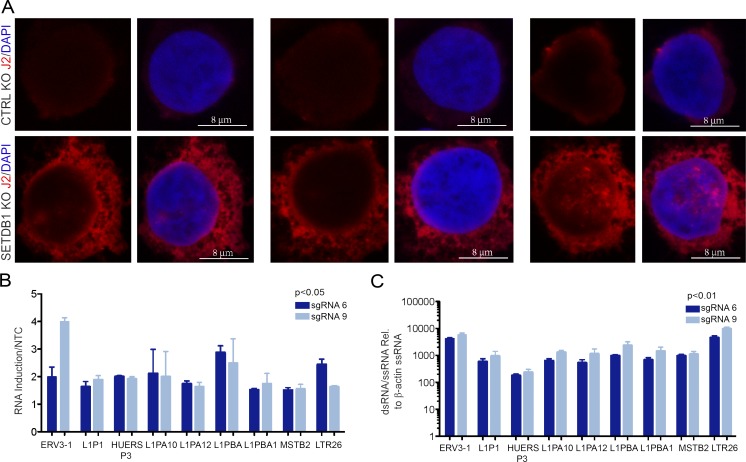
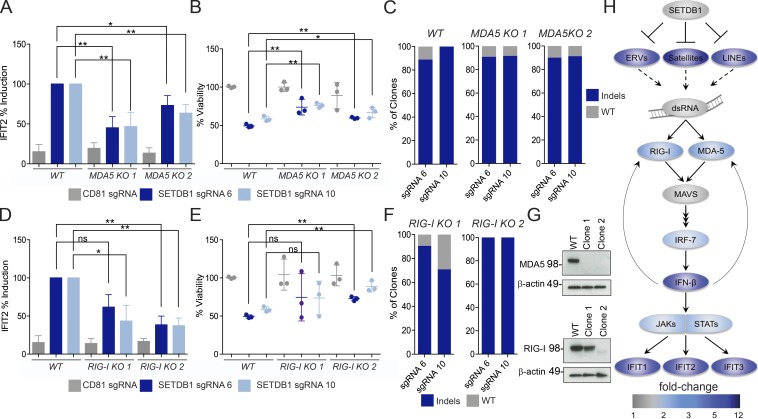
A propensity for rewiring genetic and epigenetic regulatory networks, thus enabling sustained cell proliferation, suppression of apoptosis, and the ability to evade the immune system, is vital to cancer cell propagation. An increased understanding of how this is achieved is critical for identifying or improving therapeutic interventions. In this study, using acute myeloid leukemia (AML) human cell lines and a custom CRISPR/Cas9 screening platform, we identify the H3K9 methyltransferase SETDB1 as a novel, negative regulator of innate immunity. SETDB1 is overexpressed in many cancers, and loss of this gene in AML cells triggers desilencing of retrotransposable elements that leads to the production of double-stranded RNAs (dsRNAs). This is coincident with induction of a type I interferon response and apoptosis through the dsRNA-sensing pathway. Collectively, our findings establish a unique gene regulatory axis that cancer cells can exploit to circumvent the immune system.
© 2017 Cuellar et al.
Figures
Comment in
-
Cancer cells, on your histone marks, get SETDB1, silence retrotransposons, and go!J Cell Biol. 2017 Nov 6;216(11):3429-3431. doi: 10.1083/jcb.201710068. Epub 2017 Oct 24. J Cell Biol. 2017. PMID: 29066607 Free PMC article.
References
-
- Aguirre A.J., Meyers R.M., Weir B.A., Vazquez F., Zhang C.Z., Ben-David U., Cook A., Ha G., Harrington W.F., Doshi M.B., et al. 2016. Genomic copy number dictates a gene-independent cell Response to CRISPR/Cas9 targeting. Cancer Discov. 6:914–929. 10.1158/2159-8290.CD-16-0154 - DOI - PMC - PubMed
-
- Brind’Amour J., Liu S., Hudson M., Chen C., Karimi M.M., and Lorincz M.C.. 2015. An ultra-low-input native ChIP-seq protocol for genome-wide profiling of rare cell populations. Nat. Commun. 6:6033. - PubMed
-
- Cherkasova A.P., and Selyatitskaya V.G.. 2013. Corticosteroid hormones and angiotensin-converting enzyme in the dynamics of chronic granulomatous inflammation. [In Russian]. Patol. Fiziol. Eksp. Ter. 2:26–31. - PubMed
-
- Chiappinelli K.B., Strissel P.L., Desrichard A., Li H., Henke C., Akman B., Hein A., Rote N.S., Cope L.M., Snyder A., et al. 2015. Inhibiting DNA methylation causes an interferon response in cancer via dsRNA including endogenous retroviruses. Cell. 162:974–986. (published erratum appears in Cell 2017. 169:361) 10.1016/j.cell.2015.07.011 - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
