

Dormant 5-lipoxygenase in inflammatory macrophages is triggered by exogenous arachidonic acid
- PMID: 28887514
- PMCID: PMC5591212
- DOI: 10.1038/s41598-017-11496-3
Dormant 5-lipoxygenase in inflammatory macrophages is triggered by exogenous arachidonic acid
Abstract
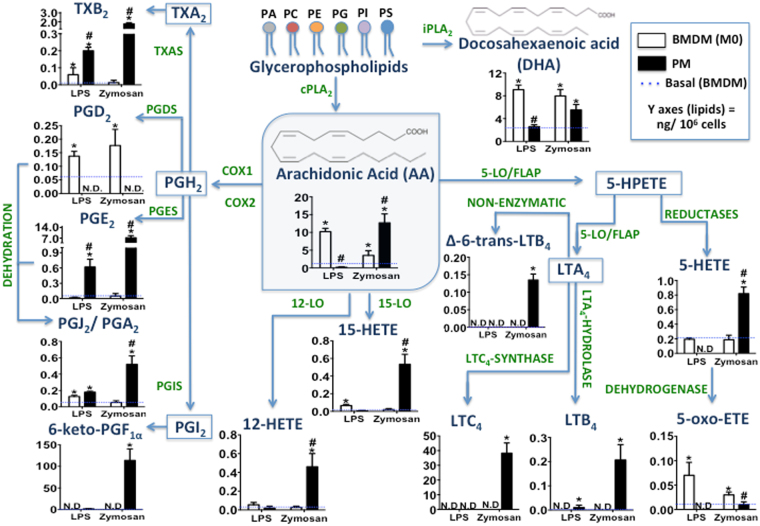
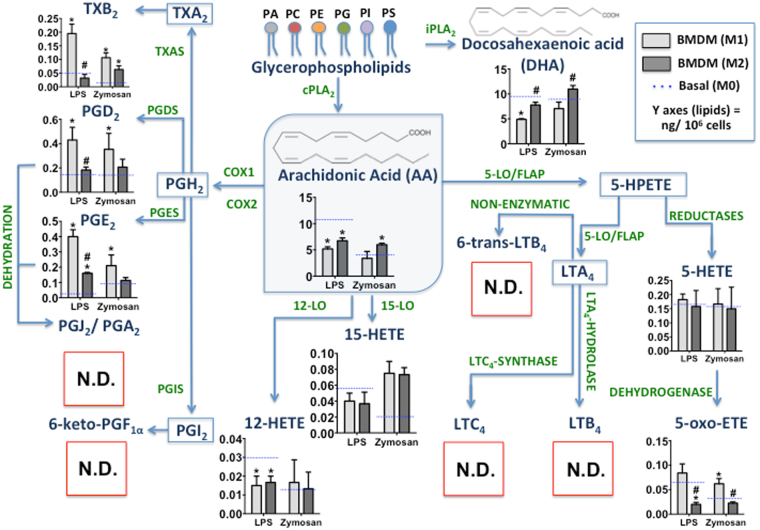
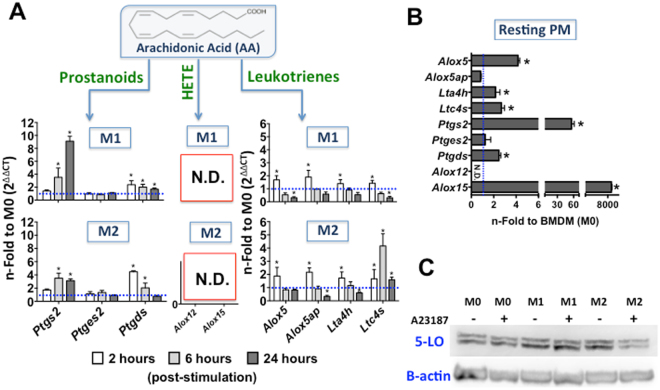
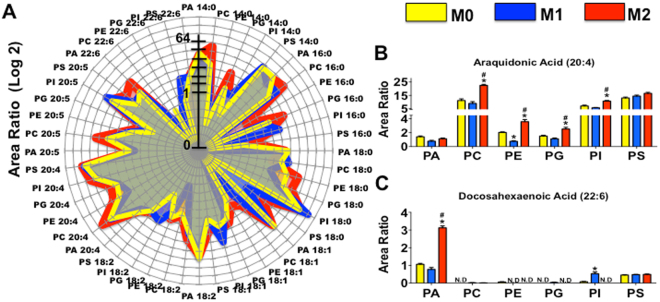
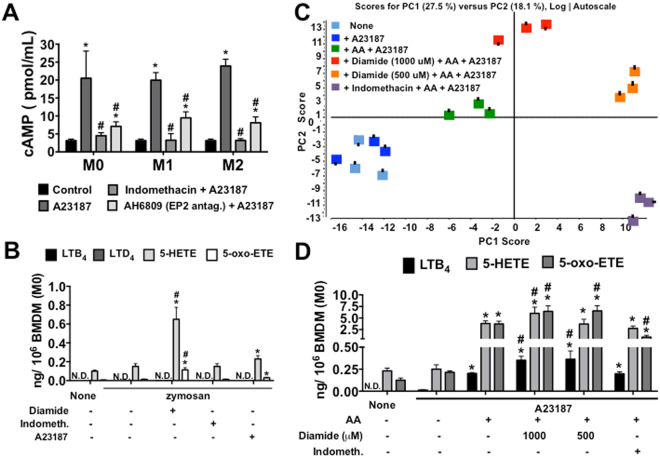
The differentiation of resident tissue macrophages from embryonic precursors and that of inflammatory macrophages from bone marrow cells leads to macrophage heterogeneity. Further plasticity is displayed through their ability to be polarized as subtypes M1 and M2 in a cell culture microenvironment. However, the detailed regulation of eicosanoid production and its involvement in macrophage biology remains unclear. Using a lipidomics approach, we demonstrated that eicosanoid production profiles between bone marrow-derived (BMDM) and peritoneal macrophages differed drastically. In polarized BMDMs, M1 and M2 phenotypes were distinguished by thromboxane B2, prostaglandin (PG) E2, and PGD2 production, in addition to lysophospholipid acyltransferase activity. Although Alox5 expression and the presence of 5-lipoxygenase (5-LO) protein in BMDMs was observed, the absence of leukotrienes production reflected an impairment in 5-LO activity, which could be triggered by addition of exogenous arachidonic acid (AA). The BMDM 5-LO regulatory mechanism was not responsive to PGE2/cAMP pathway modulation; however, treatment to reduce glutathione peroxidase activity increased 5-LO metabolite production after AA stimulation. Understanding the relationship between the eicosanoids pathway and macrophage biology may offer novel strategies for macrophage-associated disease therapy.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
