

A critical period for the trophic actions of leptin on AgRP neurons in the arcuate nucleus of the hypothalamus
- PMID: 28891045
- PMCID: PMC6040218
- DOI: 10.1002/cne.24327
A critical period for the trophic actions of leptin on AgRP neurons in the arcuate nucleus of the hypothalamus
Abstract
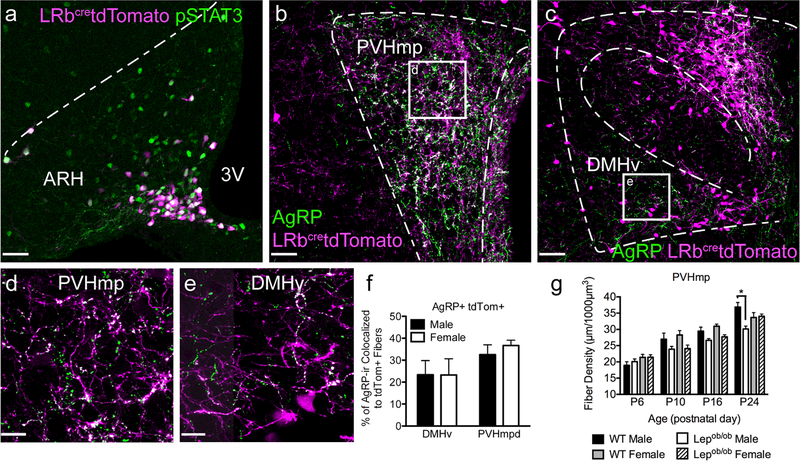
In the developing hypothalamus, the fat-derived hormone leptin stimulates the growth of axons from the arcuate nucleus of the hypothalamus (ARH) to other regions that control energy balance. These projections are significantly reduced in leptin deficient (Lepob/ob ) mice and this phenotype is largely rescued by neonatal leptin treatments. However, treatment of mature Lepob/ob mice is ineffective, suggesting that the trophic action of leptin is limited to a developmental critical period. To temporally delineate closure of this critical period for leptin-stimulated growth, we treated Lepob/ob mice with exogenous leptin during a variety of discrete time periods, and measured the density of Agouti-Related Peptide (AgRP) containing projections from the ARH to the ventral part of the dorsomedial nucleus of the hypothalamus (DMHv), and to the medial parvocellular part of the paraventricular nucleus (PVHmp). The results indicate that leptin loses its neurotrophic potential at or near postnatal day 28. The duration of leptin exposure appears to be important, with 9- or 11-day treatments found to be more effective than shorter (5-day) treatments. Furthermore, leptin treatment for 9 days or more was sufficient to restore AgRP innervation to both the PVHmp and DMHv in Lepob/ob females, but only to the DMHv in Lepob/ob males. Together, these findings reveal that the trophic actions of leptin are contingent upon timing and duration of leptin exposure, display both target and sex specificity, and that modulation of leptin-dependent circuit formation by each of these factors may carry enduring consequences for feeding behavior, metabolism, and obesity risk.
Keywords: Critical period; RRID:AB_2313908; RRID:IMSR_JAX:000632; RRID:IMSR_JAX:006417; RRID:IMSR_JAX:007914; RRID:SCR_002668; agouti-related peptide (AgRP); arcuate nucleus of the hypothalamus (ARH); leptin; sexual dimorphism.
© 2017 Wiley Periodicals, Inc.
Figures





References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
