

Structures of the human mitochondrial ribosome in native states of assembly
- PMID: 28892042
- PMCID: PMC5633077
- DOI: 10.1038/nsmb.3464
Structures of the human mitochondrial ribosome in native states of assembly
Abstract
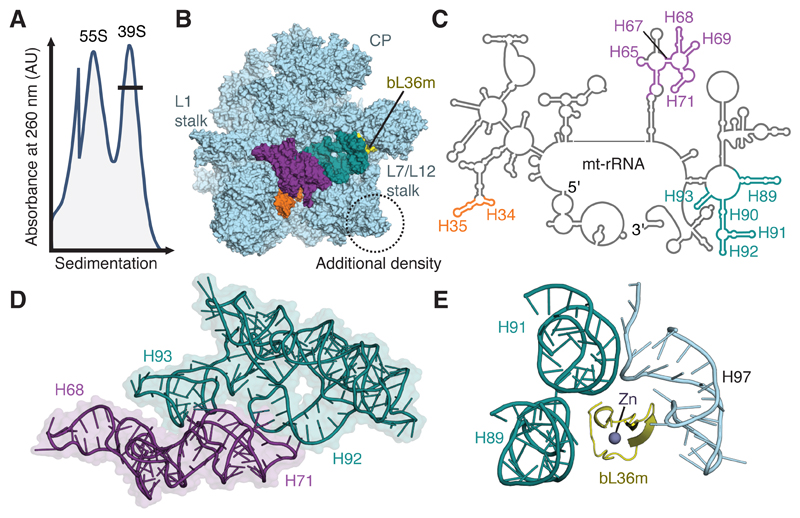
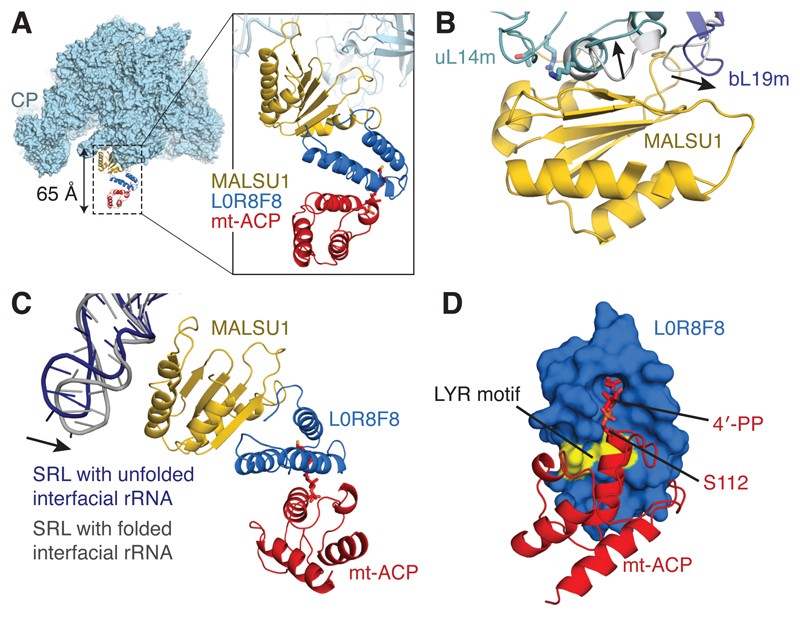
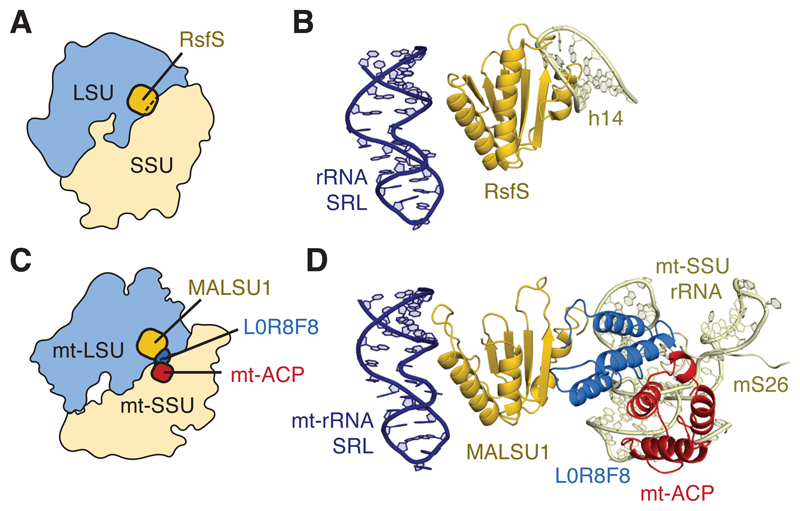
Mammalian mitochondrial ribosomes (mitoribosomes) have less rRNA content and 36 additional proteins compared with the evolutionarily related bacterial ribosome. These differences make the assembly of mitoribosomes more complex than the assembly of bacterial ribosomes, but the molecular details of mitoribosomal biogenesis remain elusive. Here, we report the structures of two late-stage assembly intermediates of the human mitoribosomal large subunit (mt-LSU) isolated from a native pool within a human cell line and solved by cryo-EM to ∼3-Å resolution. Comparison of the structures reveals insights into the timing of rRNA folding and protein incorporation during the final steps of ribosomal maturation and the evolutionary adaptations that are required to preserve biogenesis after the structural diversification of mitoribosomes. Furthermore, the structures redefine the ribosome silencing factor (RsfS) family as multifunctional biogenesis factors and identify two new assembly factors (L0R8F8 and mt-ACP) not previously implicated in mitoribosomal biogenesis.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
