

Dynamic regulation of T follicular regulatory cell responses by interleukin 2 during influenza infection
- PMID: 28892471
- PMCID: PMC5679073
- DOI: 10.1038/ni.3837
Dynamic regulation of T follicular regulatory cell responses by interleukin 2 during influenza infection
Abstract
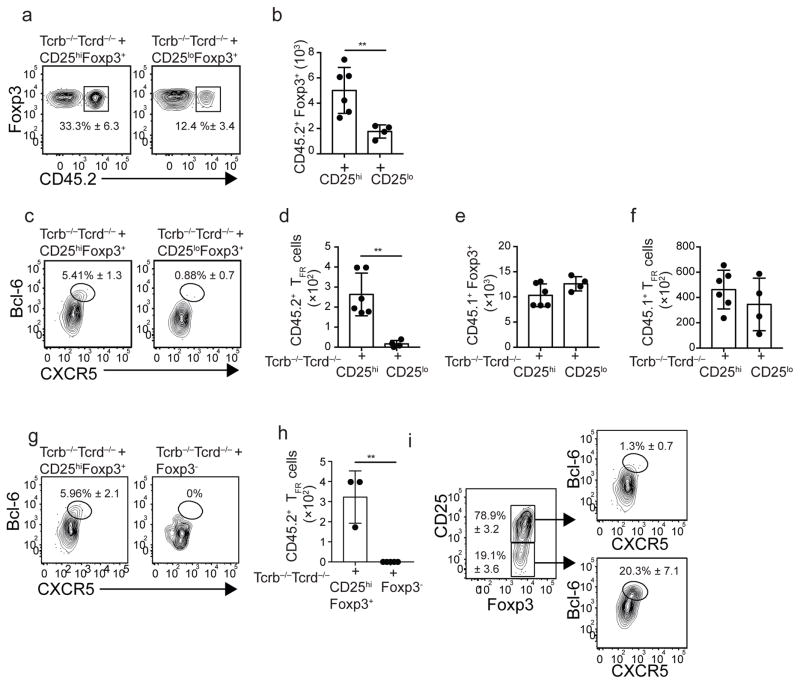
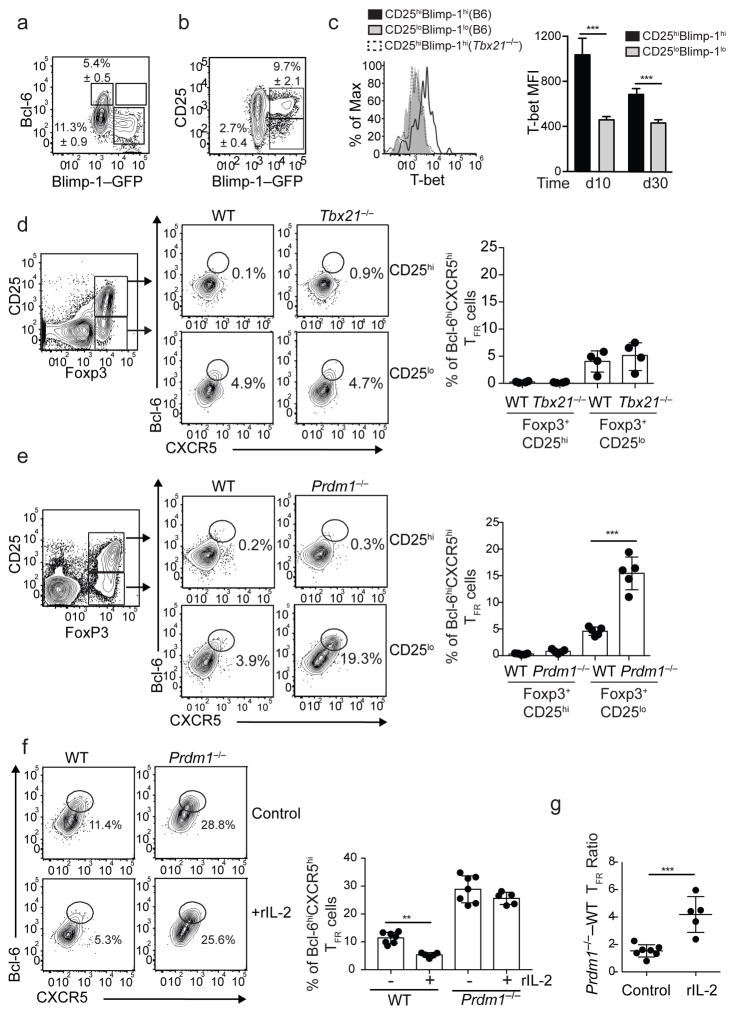
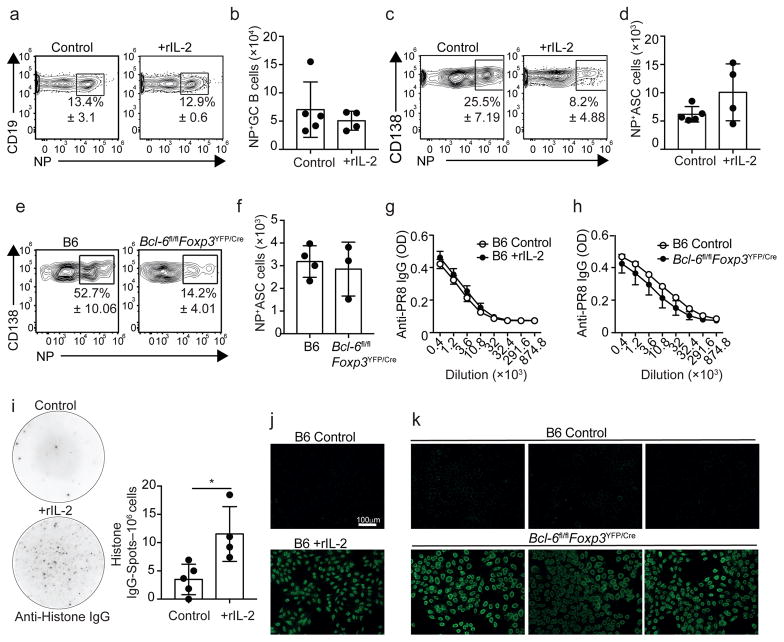
Interleukin 2 (IL-2) promotes Foxp3+ regulatory T (Treg) cell responses, but inhibits T follicular helper (TFH) cell development. However, it is not clear how IL-2 affects T follicular regulatory (TFR) cells, a cell type with properties of both Treg and TFH cells. Using an influenza infection model, we found that high IL-2 concentrations at the peak of the infection prevented TFR cell development by a Blimp-1-dependent mechanism. However, once the immune response resolved, some Treg cells downregulated CD25, upregulated Bcl-6 and differentiated into TFR cells, which then migrated into the B cell follicles to prevent the expansion of self-reactive B cell clones. Thus, unlike its effects on conventional Treg cells, IL-2 inhibits TFR cell responses.
Figures





Comment in
-
TFR cells trump autoimmune antibody responses to limit sedition.Nat Immunol. 2017 Oct 18;18(11):1185-1186. doi: 10.1038/ni.3856. Nat Immunol. 2017. PMID: 29044242 No abstract available.
-
Regulatory T Cells: The IL-2 gauge.Nat Rev Immunol. 2017 Oct 27;17(11):662-663. doi: 10.1038/nri.2017.123. Nat Rev Immunol. 2017. PMID: 29076497 No abstract available.
References
-
- Burchill MA, Yang J, Vogtenhuber C, Blazar BR, Farrar MA. IL-2 receptor beta-dependent STAT5 activation is required for the development of Foxp3+ regulatory T cells. J Immunol. 2007;178:280–290. - PubMed
-
- Fontenot JD, Rasmussen JP, Gavin MA, Rudensky AY. A function for interleukin 2 in Foxp3-expressing regulatory T cells. Nature immunology. 2005;6:1142–1151. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
