

Functional Amyloid Protection in the Eye Lens: Retention of α-Crystallin Molecular Chaperone Activity after Modification into Amyloid Fibrils
- PMID: 28895938
- PMCID: PMC5618248
- DOI: 10.3390/biom7030067
Functional Amyloid Protection in the Eye Lens: Retention of α-Crystallin Molecular Chaperone Activity after Modification into Amyloid Fibrils
Abstract
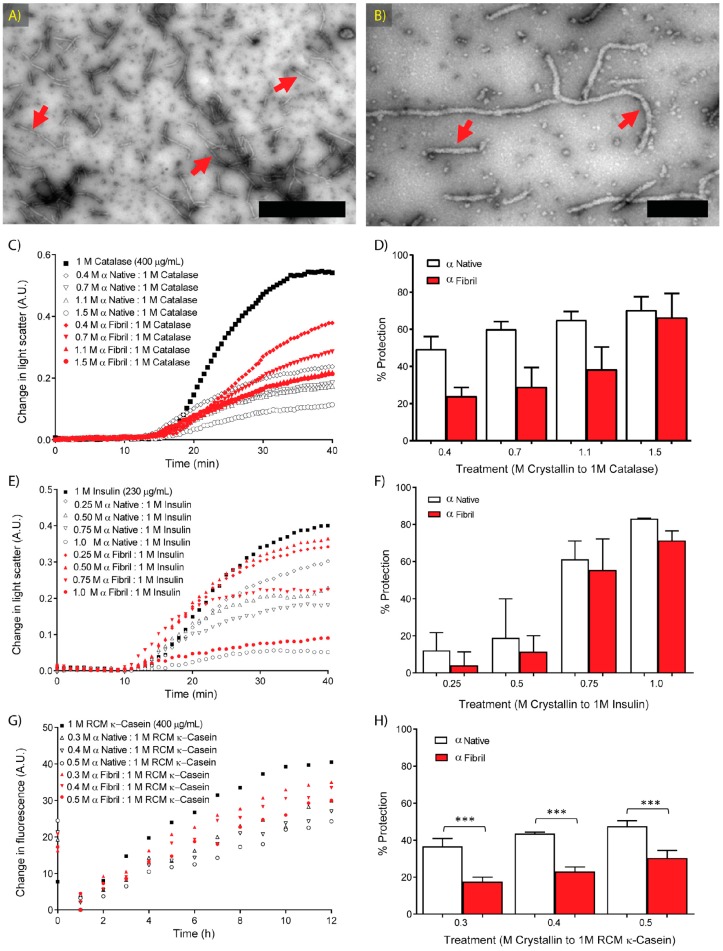
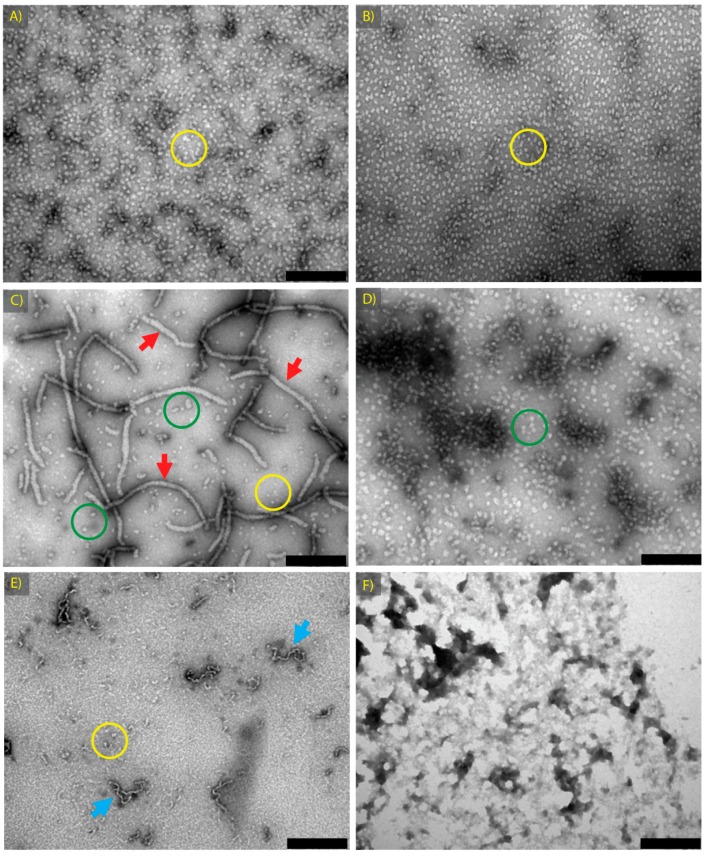
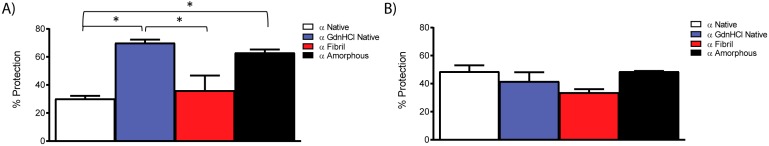
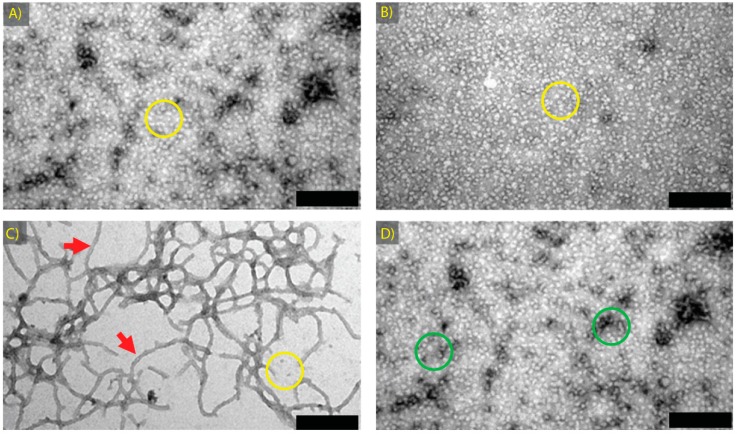
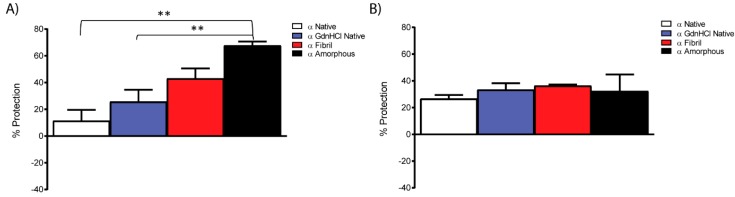
Amyloid fibril formation occurs from a wide range of peptides and proteins and is typically associated with a loss of protein function and/or a gain of toxic function, as the native structure of the protein undergoes major alteration to form a cross β-sheet array. It is now well recognised that some amyloid fibrils have a biological function, which has led to increased interest in the potential that these so-called functional amyloids may either retain the function of the native protein, or gain function upon adopting a fibrillar structure. Herein, we investigate the molecular chaperone ability of α-crystallin, the predominant eye lens protein which is composed of two related subunits αA- and αB-crystallin, and its capacity to retain and even enhance its chaperone activity after forming aggregate structures under conditions of thermal and chemical stress. We demonstrate that both eye lens α-crystallin and αB-crystallin (which is also found extensively outside the lens) retain, to a significant degree, their molecular chaperone activity under conditions of structural change, including after formation into amyloid fibrils and amorphous aggregates. The results can be related directly to the effects of aging on the structure and chaperone function of α-crystallin in the eye lens, particularly its ability to prevent crystallin protein aggregation and hence lens opacification associated with cataract formation.
Keywords: amyloid fibril; molecular chaperone; protein aggregation; protein unfolding; small heat-shock protein.
Conflict of interest statement
The authors declare no conflict of interest. The funding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
