

Rebuilding Chromosomes After Catastrophe: Emerging Mechanisms of Chromothripsis
- PMID: 28899600
- PMCID: PMC5696049
- DOI: 10.1016/j.tcb.2017.08.005
Rebuilding Chromosomes After Catastrophe: Emerging Mechanisms of Chromothripsis
Abstract
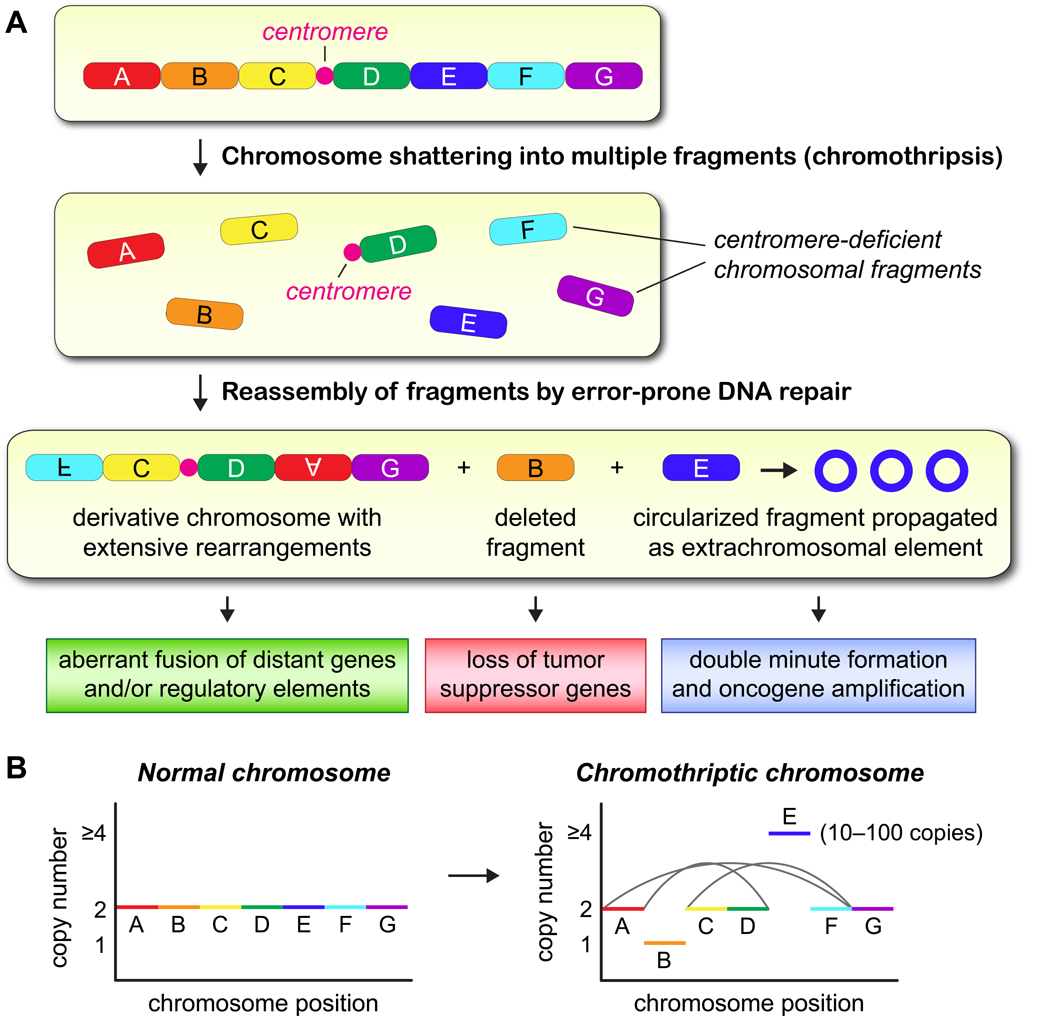
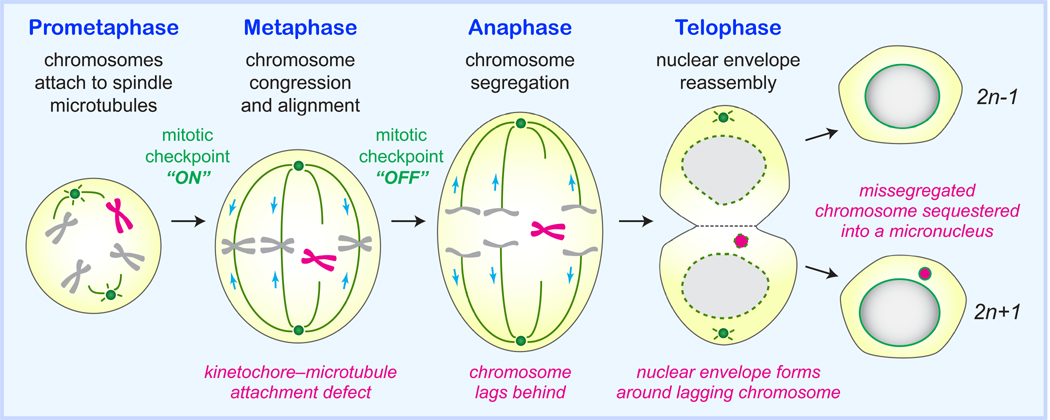
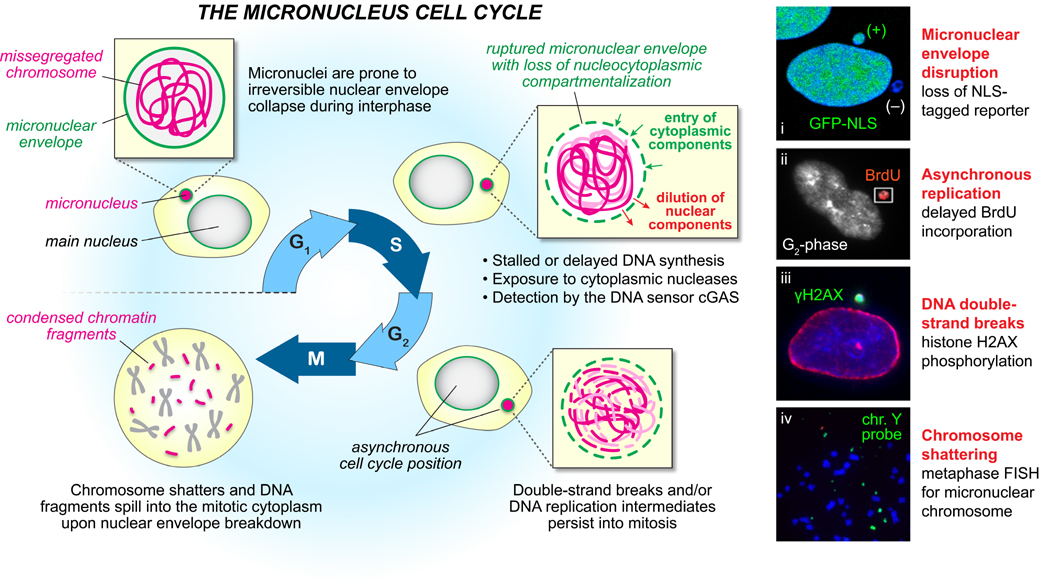
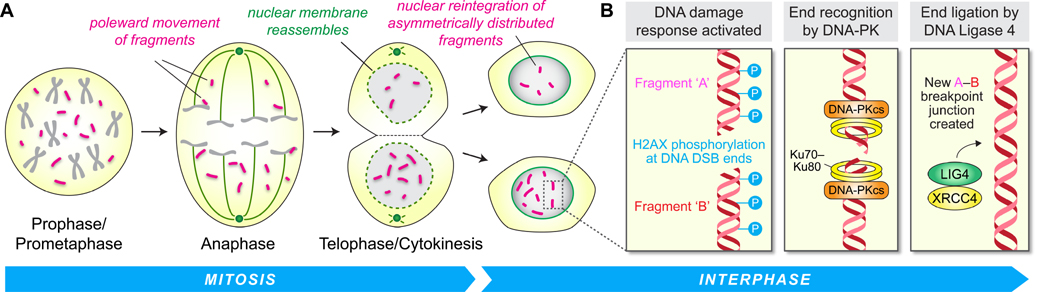
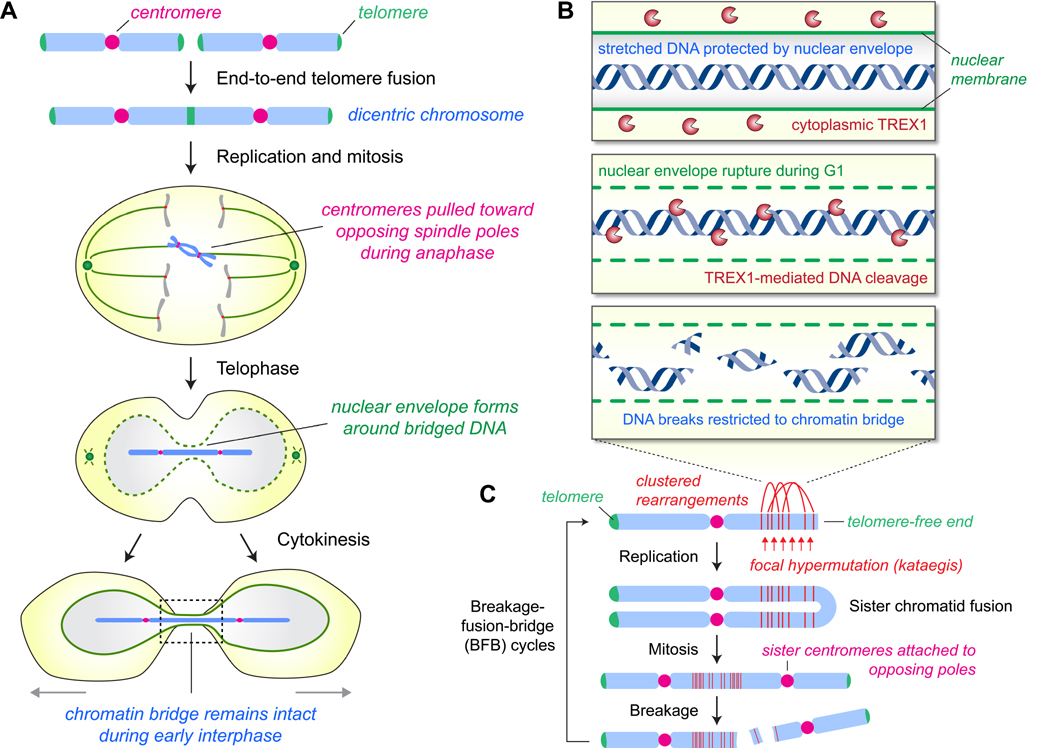
Cancer genome sequencing has identified chromothripsis, a complex class of structural genomic rearrangements involving the apparent shattering of an individual chromosome into tens to hundreds of fragments. An initial error during mitosis, producing either chromosome mis-segregation into a micronucleus or chromatin bridge interconnecting two daughter cells, can trigger the catastrophic pulverization of the spatially isolated chromosome. The resultant chromosomal fragments are religated in random order by DNA double-strand break repair during the subsequent interphase. Chromothripsis scars the cancer genome with localized DNA rearrangements that frequently generate extensive copy number alterations, oncogenic gene fusion products, and/or tumor suppressor gene inactivation. Here we review emerging mechanisms underlying chromothripsis with a focus on the contribution of cell division errors caused by centromere dysfunction.
Keywords: DNA repair; chromosome rearrangements; chromothripsis; genomic instability; micronuclei; mitosis.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
References
-
- Nowell PC, Hungerford DA. Minute Chromosome in Human Chronic Granulocytic Leukemia. Science. 1960;132:1497–1497.
-
- Druker BJ, Talpaz M, Resta DJ, Peng B, Buchdunger E, Ford JM, Lydon NB, Kantarjian H, Capdeville R, Ohno-Jones S, et al. Efficacy and safety of a specific inhibitor of the BCR-ABL tyrosine kinase in chronic myeloid leukemia. N Engl J Med. 2001;344:1031–1037. - PubMed
-
- Fearon ER, Vogelstein B. A genetic model for colorectal tumorigenesis. Cell. 1990;61:759–767. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
