

Adolescent THC Exposure Causes Enduring Prefrontal Cortical Disruption of GABAergic Inhibition and Dysregulation of Sub-Cortical Dopamine Function
- PMID: 28900286
- PMCID: PMC5595795
- DOI: 10.1038/s41598-017-11645-8
Adolescent THC Exposure Causes Enduring Prefrontal Cortical Disruption of GABAergic Inhibition and Dysregulation of Sub-Cortical Dopamine Function
Abstract
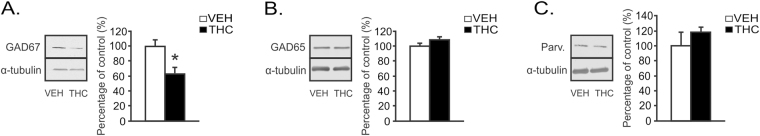
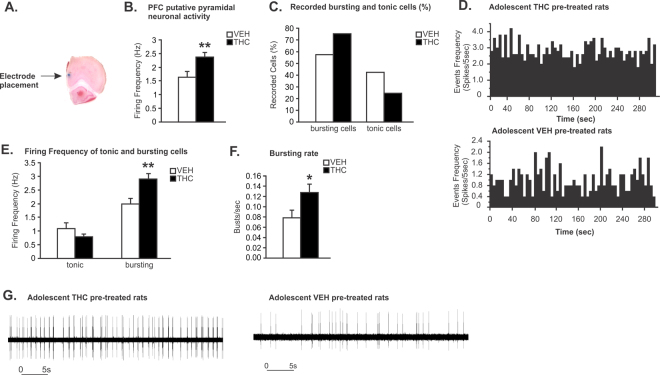
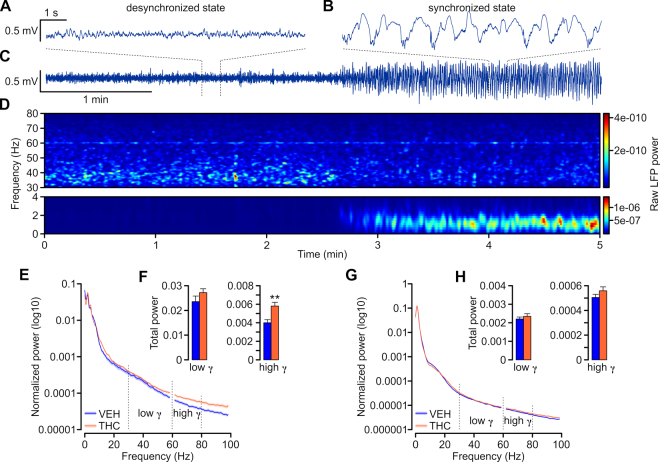
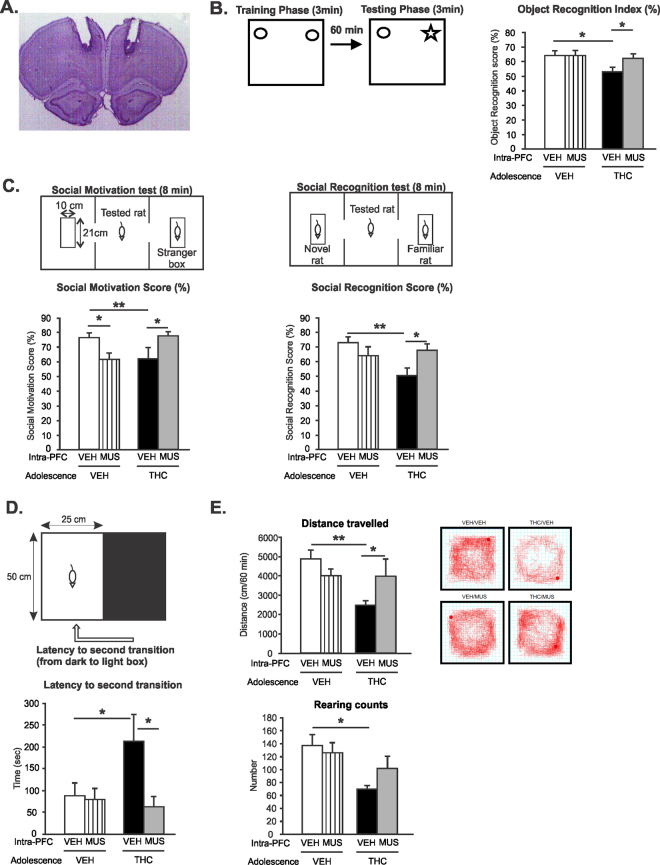
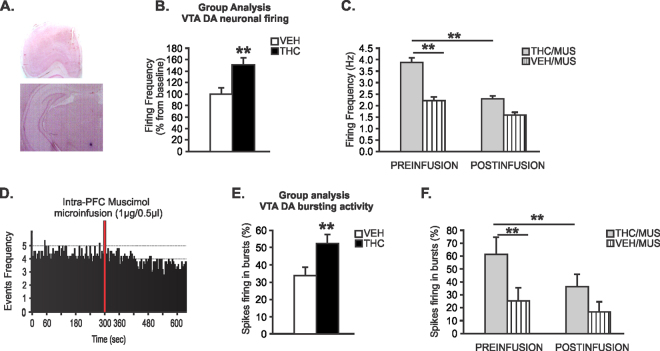
Chronic adolescent marijuana use has been linked to the later development of psychiatric diseases such as schizophrenia. GABAergic hypofunction in the prefrontal cortex (PFC) is a cardinal pathological feature of schizophrenia and may be a mechanism by which the PFC loses its ability to regulate sub-cortical dopamine (DA) resulting in schizophrenia-like neuropsychopathology. In the present study, we exposed adolescent rats to Δ-9-tetra-hydrocannabinol (THC), the psychoactive component in marijuana. At adulthood, we characterized the functionality of PFC GABAergic neurotransmission and its regulation of sub-cortical DA function using molecular, behavioral and in-vivo electrophysiological analyses. Our findings revealed a persistent attenuation of PFC GABAergic function combined with a hyperactive neuronal state in PFC neurons and associated disruptions in cortical gamma oscillatory activity. These PFC abnormalities were accompanied by hyperactive DAergic neuronal activity in the ventral tegmental area (VTA) and behavioral and cognitive abnormalities similar to those observed in psychiatric disorders. Remarkably, these neuronal and behavioral effects were reversed by pharmacological activation of GABAA receptors in the PFC. Together, these results identify a mechanistic link between dysregulated frontal cortical GABAergic inhibition and sub-cortical DAergic dysregulation, characteristic of well-established neuropsychiatric endophenotypes.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
