

New tricks for the glycyl radical enzyme family
- PMID: 28901199
- PMCID: PMC5911432
- DOI: 10.1080/10409238.2017.1373741
New tricks for the glycyl radical enzyme family
Abstract
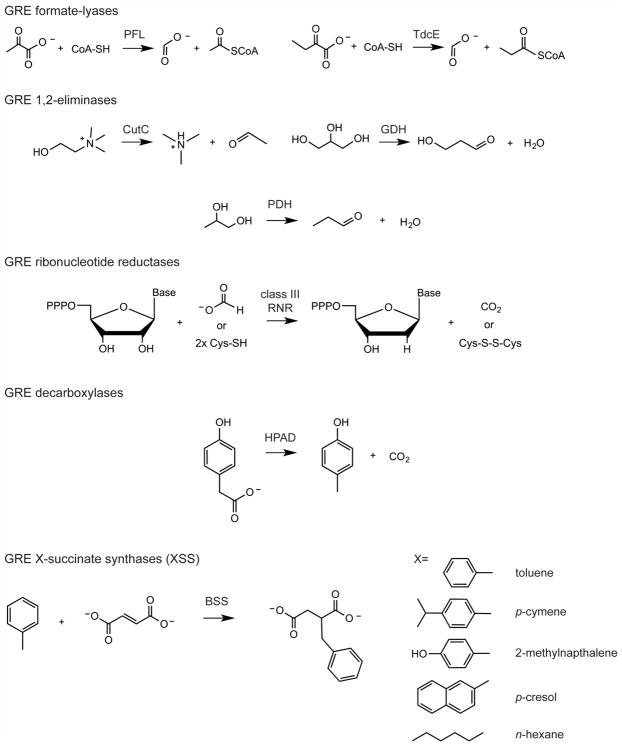
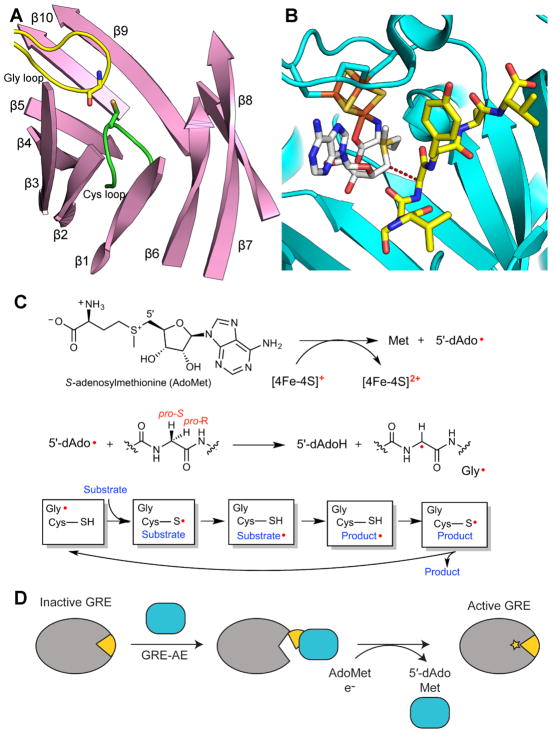
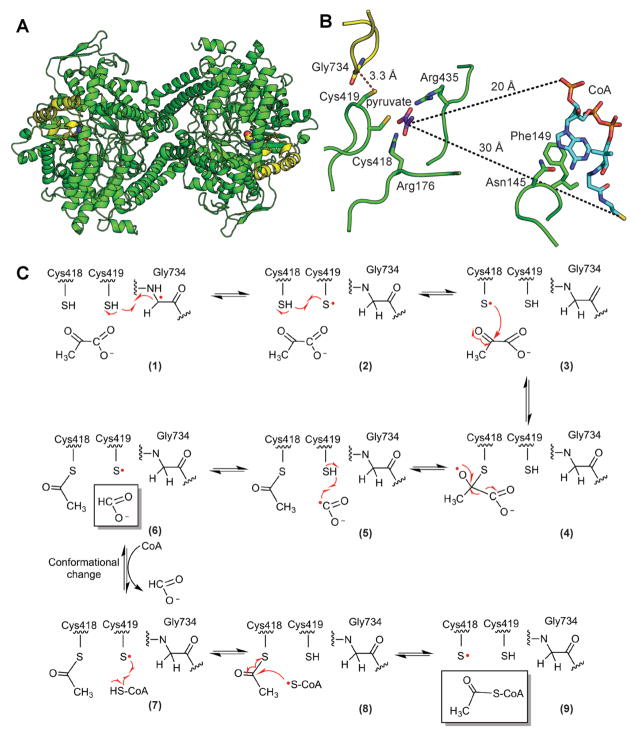
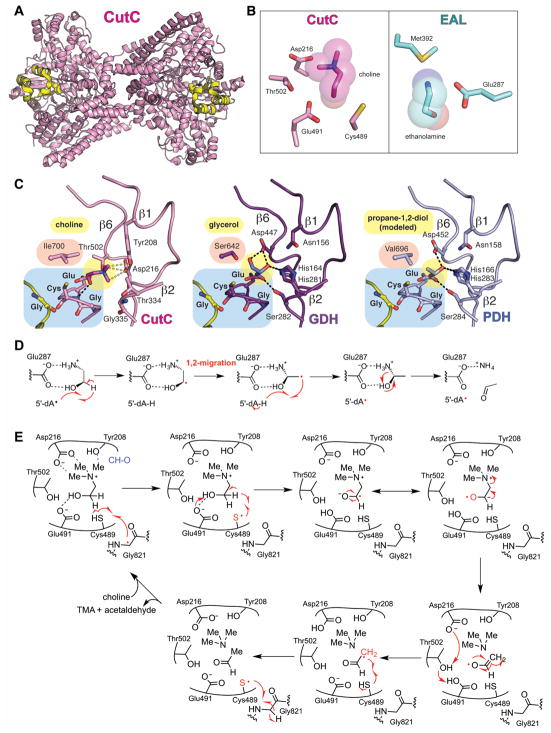
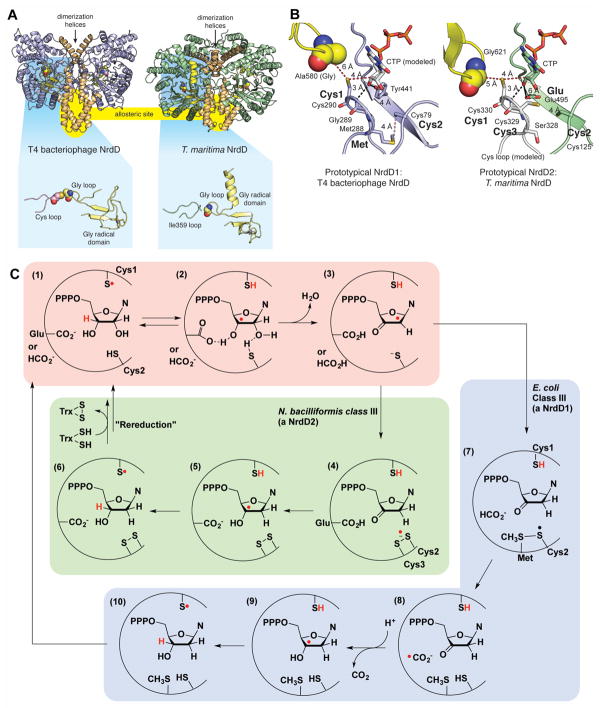
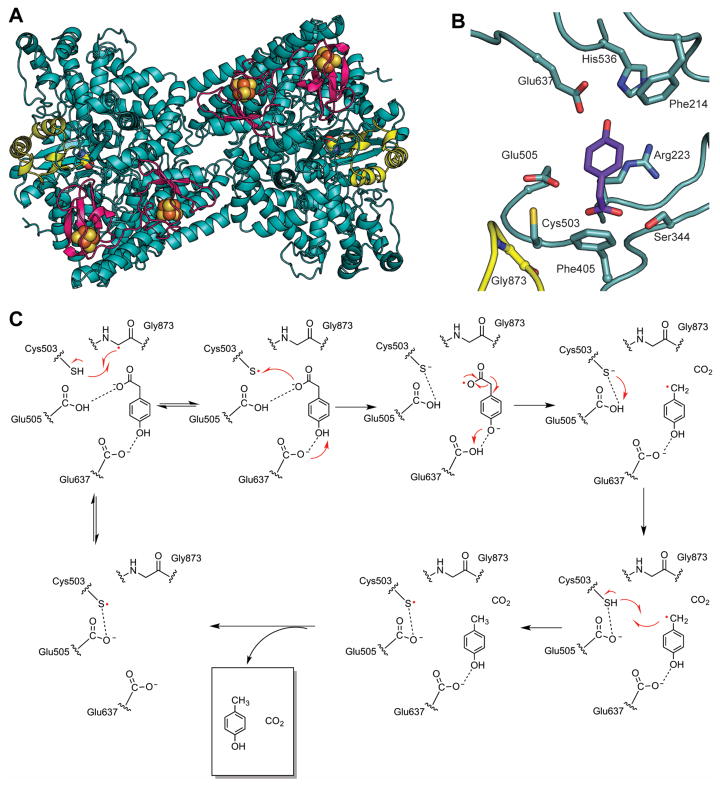
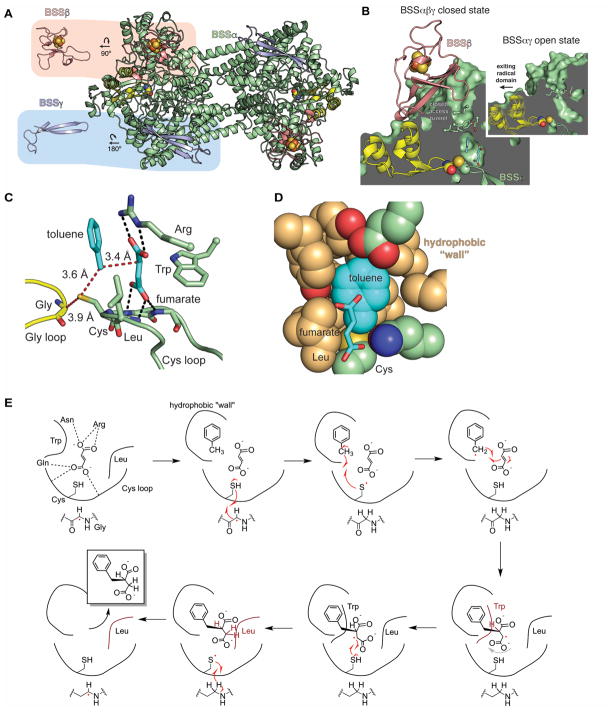
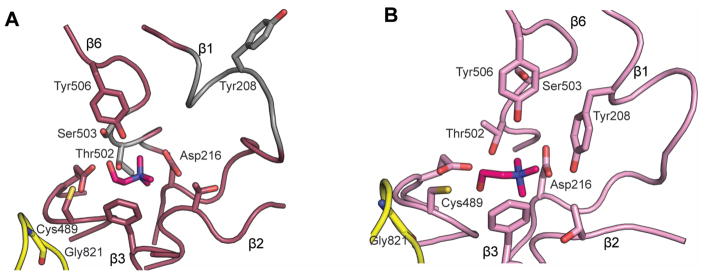
Glycyl radical enzymes (GREs) are important biological catalysts in both strict and facultative anaerobes, playing key roles both in the human microbiota and in the environment. GREs contain a backbone glycyl radical that is post-translationally installed, enabling radical-based mechanisms. GREs function in several metabolic pathways including mixed acid fermentation, ribonucleotide reduction and the anaerobic breakdown of the nutrient choline and the pollutant toluene. By generating a substrate-based radical species within the active site, GREs enable C-C, C-O and C-N bond breaking and formation steps that are otherwise challenging for nonradical enzymes. Identification of previously unknown family members from genomic data and the determination of structures of well-characterized GREs have expanded the scope of GRE-catalyzed reactions as well as defined key features that enable radical catalysis. Here, we review the structures and mechanisms of characterized GREs, classifying members into five categories. We consider the open questions about each of the five GRE classes and evaluate the tools available to interrogate uncharacterized GREs.
Keywords: Glycyl radical enzymes; anaerobic metabolism; benzylsuccinate synthase; choline trimethylamine-lyase; class III ribonucleotide reductase; pyruvate formate-lyase; radical chemistry; radical decarboxylases.
Conflict of interest statement
The authors declare they have no conflicts of interest. This work was supported in part by the National Institutes of Health (NIH) GM069857 (C.L.D.), the NIH Pre-Doctoral Training Grant T32GM007287 (C.D.D.), the National Science Foundation (NSF) Graduate Research Fellowship under Grant No. 1122374 (L.R.F.B.) and 0645960 (M.A.F.). C.L.D is a Howard Hughes Medical Institute (HHMI) Investigator.
Figures
Similar articles
-
A stable organic free radical in anaerobic benzylsuccinate synthase of Azoarcus sp. strain T.J Biol Chem. 2001 Apr 20;276(16):12924-7. doi: 10.1074/jbc.M009453200. Epub 2001 Jan 30. J Biol Chem. 2001. PMID: 11278506
-
New glycyl radical enzymes catalysing key metabolic steps in anaerobic bacteria.Biol Chem. 2005 Oct;386(10):981-8. doi: 10.1515/BC.2005.114. Biol Chem. 2005. PMID: 16218870 Review.
-
The Autonomous Glycyl Radical Protein GrcA Restores Activity to Inactive Full-Length Pyruvate Formate-Lyase In Vivo.J Bacteriol. 2022 May 17;204(5):e0007022. doi: 10.1128/jb.00070-22. Epub 2022 Apr 4. J Bacteriol. 2022. PMID: 35377165 Free PMC article.
-
A glycyl radical site in the crystal structure of a class III ribonucleotide reductase.Science. 1999 Mar 5;283(5407):1499-504. doi: 10.1126/science.283.5407.1499. Science. 1999. PMID: 10066165
-
Glycyl radical activating enzymes: structure, mechanism, and substrate interactions.Arch Biochem Biophys. 2014 Mar 15;546:64-71. doi: 10.1016/j.abb.2014.01.020. Epub 2014 Jan 31. Arch Biochem Biophys. 2014. PMID: 24486374 Free PMC article. Review.
Cited by
-
Peripheral Methionine Residues Impact Flavin Photoreduction and Protonation in an Engineered LOV Domain Light Sensor.Biochemistry. 2021 Apr 20;60(15):1148-1164. doi: 10.1021/acs.biochem.1c00064. Epub 2021 Mar 31. Biochemistry. 2021. PMID: 33787242 Free PMC article.
-
Two radical-dependent mechanisms for anaerobic degradation of the globally abundant organosulfur compound dihydroxypropanesulfonate.Proc Natl Acad Sci U S A. 2020 Jul 7;117(27):15599-15608. doi: 10.1073/pnas.2003434117. Epub 2020 Jun 22. Proc Natl Acad Sci U S A. 2020. PMID: 32571930 Free PMC article.
-
Nucleotide binding to the ATP-cone in anaerobic ribonucleotide reductases allosterically regulates activity by modulating substrate binding.Elife. 2024 Jul 5;12:RP89292. doi: 10.7554/eLife.89292. Elife. 2024. PMID: 38968292 Free PMC article.
-
Selenocysteine Substitution in a Class I Ribonucleotide Reductase.Biochemistry. 2019 Dec 17;58(50):5074-5084. doi: 10.1021/acs.biochem.9b00973. Epub 2019 Dec 6. Biochemistry. 2019. PMID: 31774661 Free PMC article.
-
Mechanistic Studies of a Skatole-Forming Glycyl Radical Enzyme Suggest Reaction Initiation via Hydrogen Atom Transfer.J Am Chem Soc. 2022 Jun 29;144(25):11110-11119. doi: 10.1021/jacs.1c13580. Epub 2022 Jun 15. J Am Chem Soc. 2022. PMID: 35704859 Free PMC article.
References
-
- Adhikari U, Scheiner S. Magnitude and mechanism of charge enhancement of CH··O hydrogen bonds. J Phys Chem A. 2013;117:10551–62. - PubMed
-
- Andersson J, Westman M, Hofer A, Sjoberg BM. Allosteric regulation of the class III anaerobic ribonucleotide reductase from bacteriophage T4. J Biol Chem. 2000;275:19443–8. - PubMed
-
- Aurelius O, Johansson R, Bagenholm V, Lundin D, Tholander F, Balhuizen A, Beck T, Sahlin M, Sjoberg BM, Mulliez E, Logan DT. The Crystal Structure of Thermotoga maritima Class III Ribonucleotide Reductase Lacks a Radical Cysteine Pre-Positioned in the Active Site. PLoS One. 2015;10:e0128199. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources