

In planta expression of hyperthermophilic enzymes as a strategy for accelerated lignocellulosic digestion
- PMID: 28904370
- PMCID: PMC5597601
- DOI: 10.1038/s41598-017-11026-1
In planta expression of hyperthermophilic enzymes as a strategy for accelerated lignocellulosic digestion
Abstract
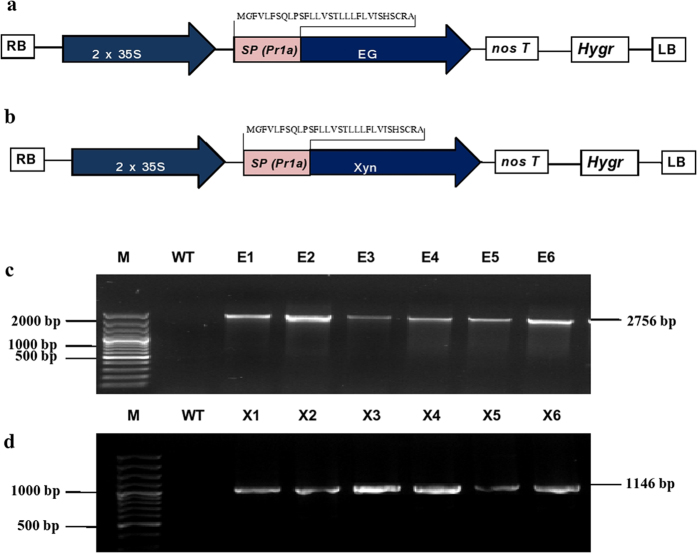
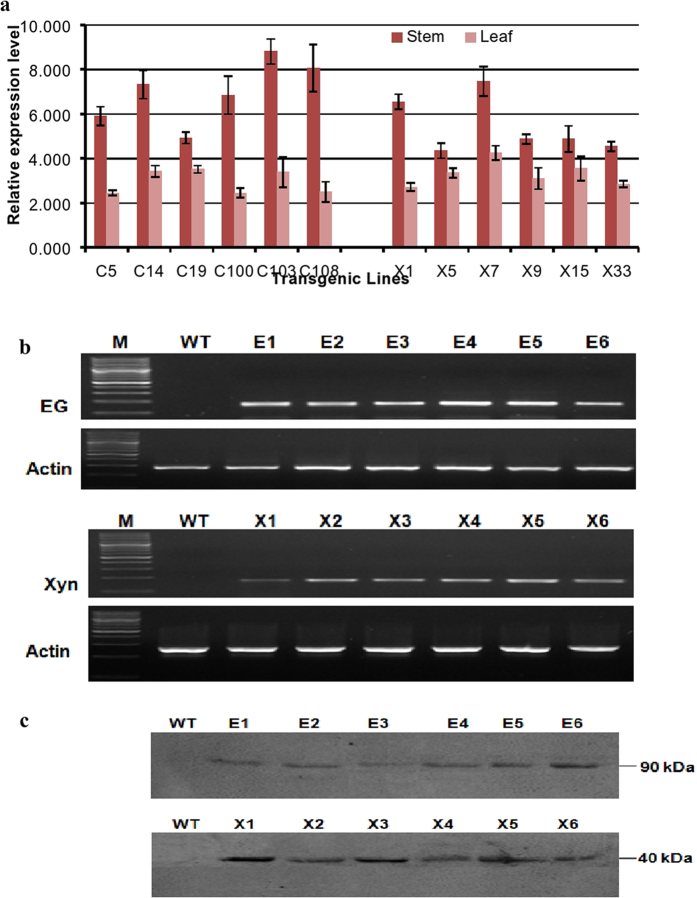
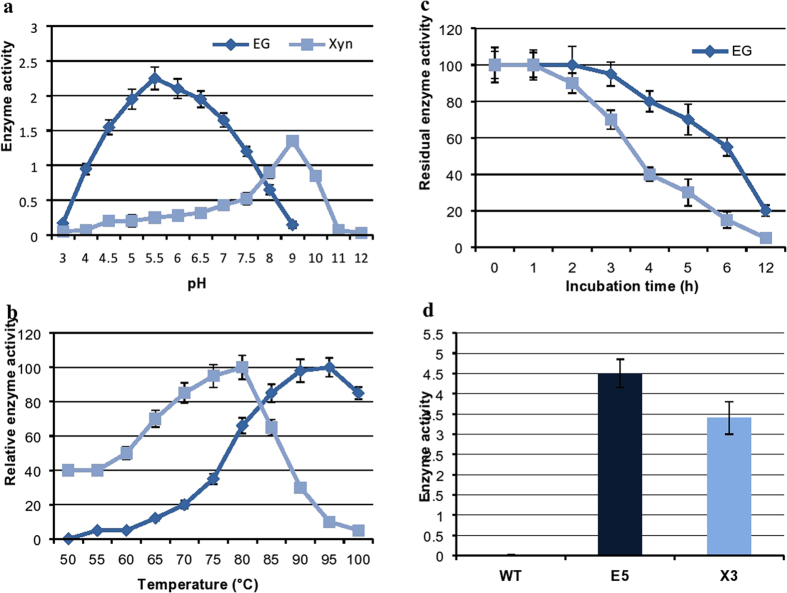
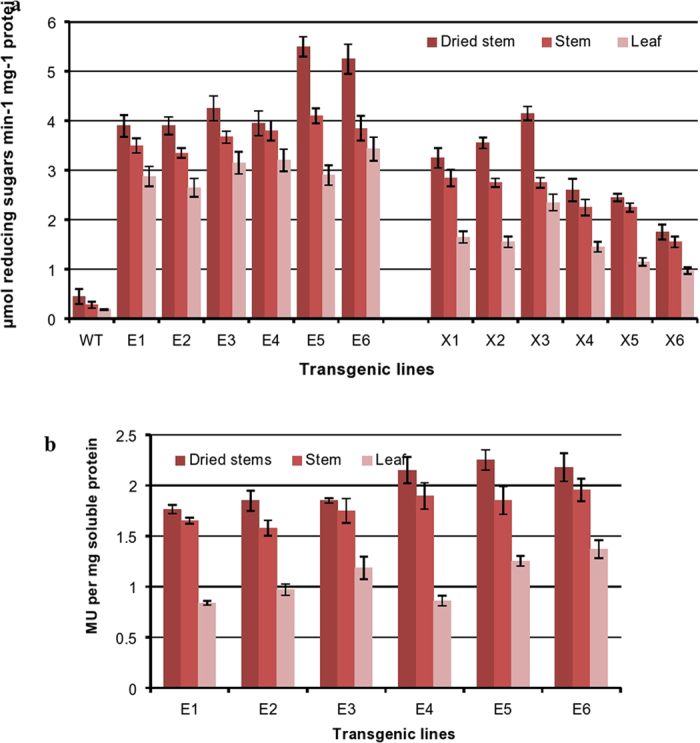
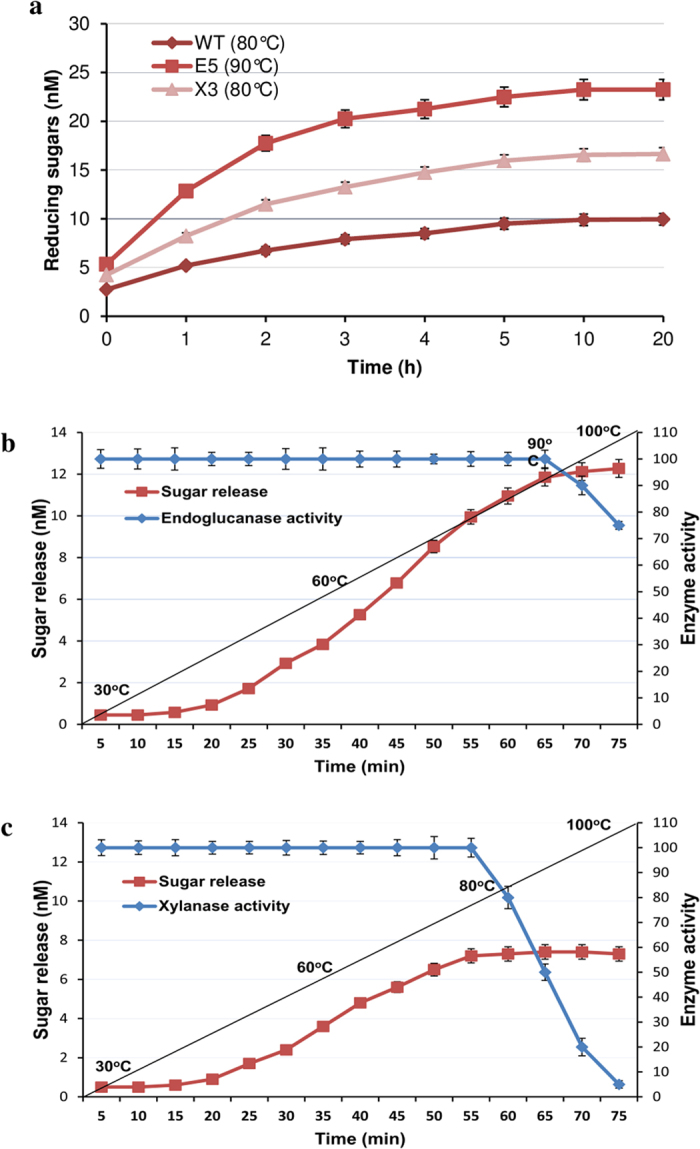
Conversion of lignocellulosic biomass to biofuels and biomaterials suffers from high production costs associated with biomass pretreatment and enzymatic hydrolysis. In-planta expression of lignocellulose-digesting enzymes is a promising approach to reduce these cost elements. However, this approach faces a number of challenges, including auto-hydrolysis of developing cell walls, plant growth and yield penalties, low expression levels and the limited stability of expressed enzymes at the high temperatures generally used for biomass processing to release fermentable sugars. To overcome these challenges we expressed codon-optimized recombinant hyperthermophilic endoglucanase (EG) and xylanase (Xyn) genes in A. thaliana. Transgenic Arabidopsis lines expressing EG and Xyn enzymes at high levels without any obvious plant growth or yield penalties were selected for further analysis. The highest enzyme activities were observed in the dry stems of transgenic lines, indicating that the enzymes were not degraded during stem senescence and storage. Biomass from transgenic lines exhibited improved saccharification efficiency relative to WT control plants. We conclude that the expression of hyperthermophilic enzymes in plants is a promising approach for combining pretreatment and enzymatic hydrolysis processes in lignocellulosic digestion. This study provides a valid foundation for further studies involving in planta co-expression of core and accessory lignocellulose-digesting enzymes.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
