

Rapid Discovery of De Novo Deleterious Mutations in Cattle Enhances the Value of Livestock as Model Species
- PMID: 28904385
- PMCID: PMC5597596
- DOI: 10.1038/s41598-017-11523-3
Rapid Discovery of De Novo Deleterious Mutations in Cattle Enhances the Value of Livestock as Model Species
Abstract
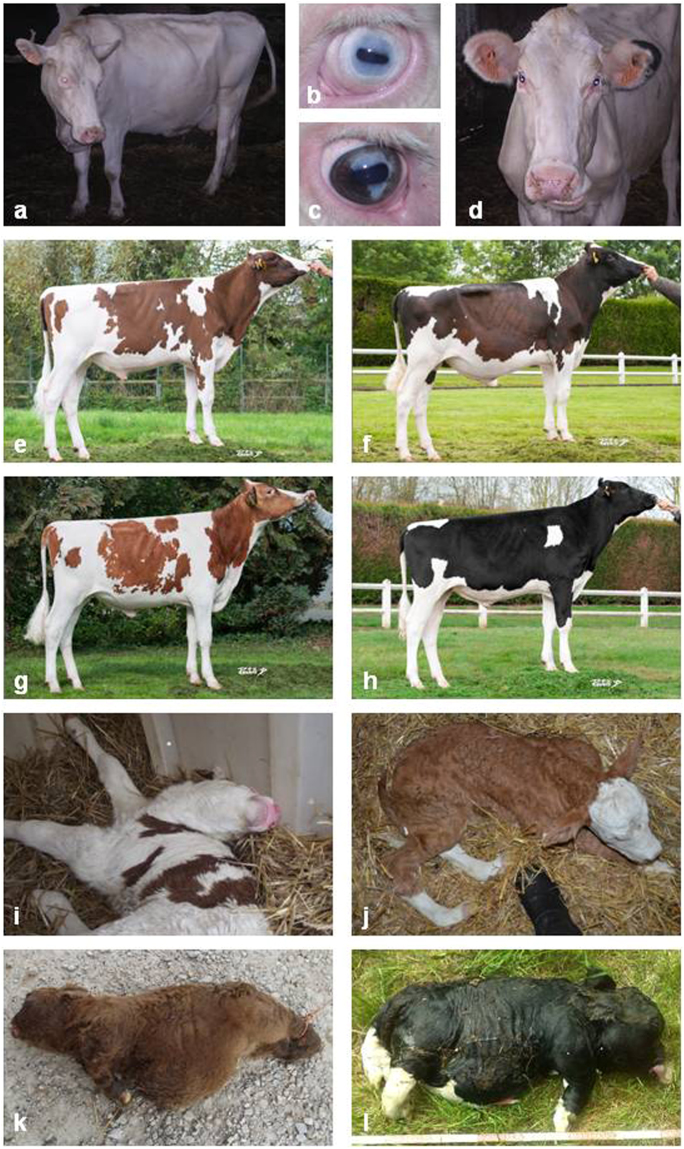
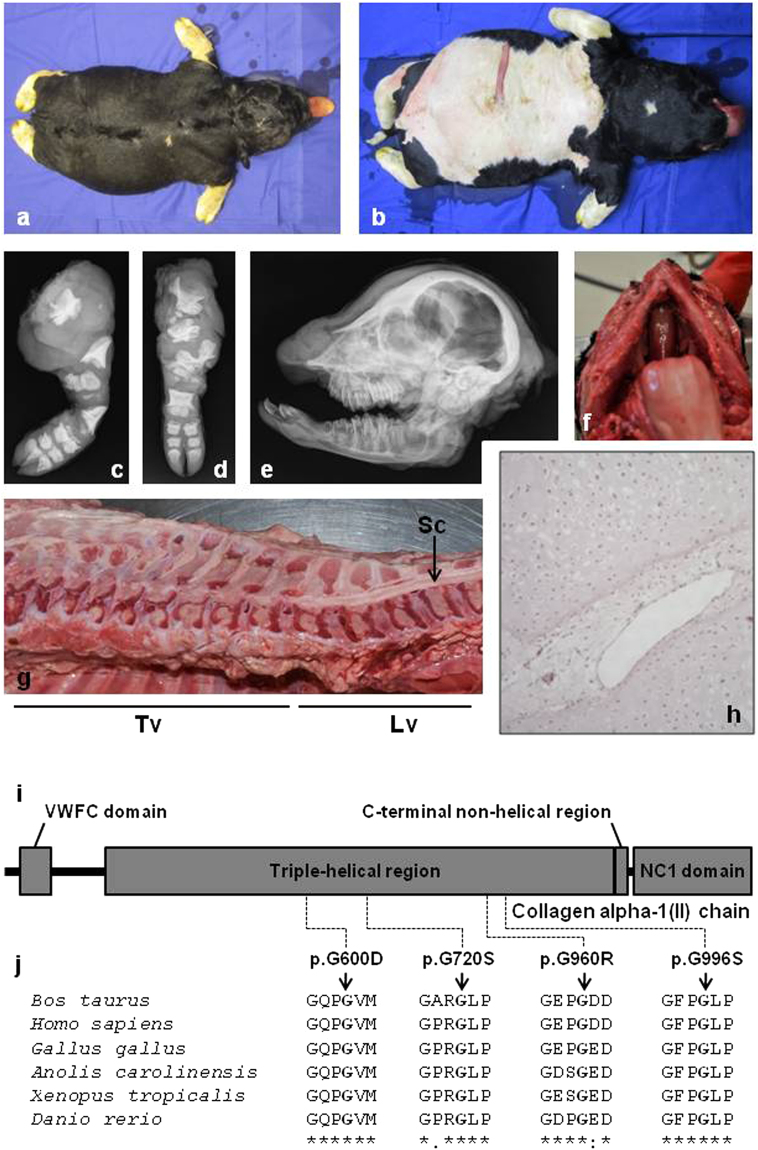
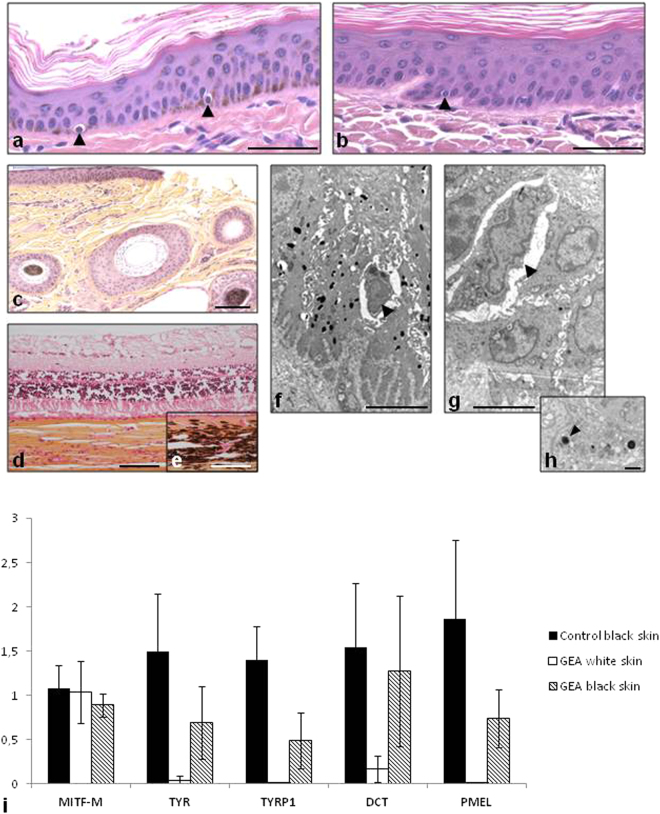
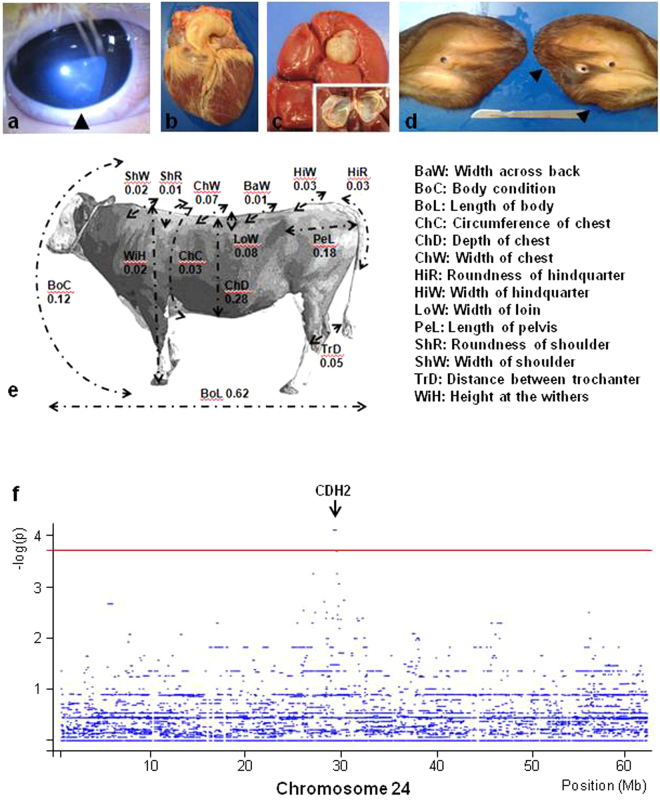
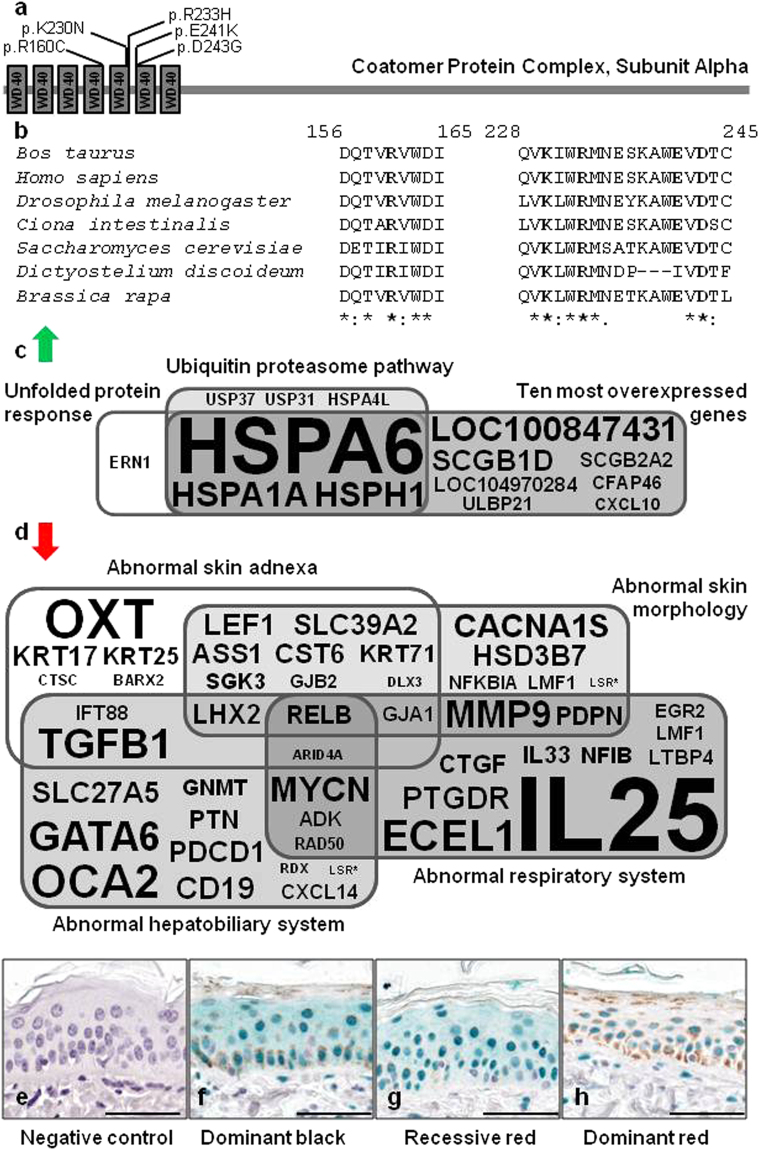
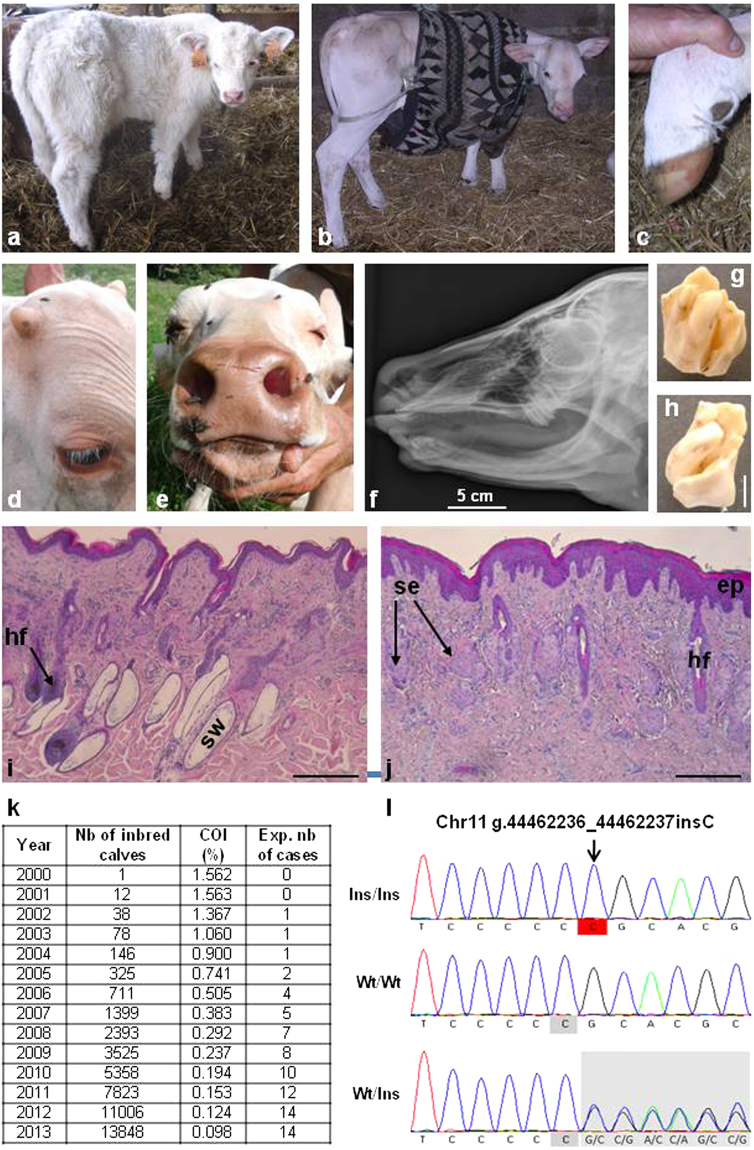
In humans, the clinical and molecular characterization of sporadic syndromes is often hindered by the small number of patients and the difficulty in developing animal models for severe dominant conditions. Here we show that the availability of large data sets of whole-genome sequences, high-density SNP chip genotypes and extensive recording of phenotype offers an unprecedented opportunity to quickly dissect the genetic architecture of severe dominant conditions in livestock. We report on the identification of seven dominant de novo mutations in CHD7, COL1A1, COL2A1, COPA, and MITF and exploit the structure of cattle populations to describe their clinical consequences and map modifier loci. Moreover, we demonstrate that the emergence of recessive genetic defects can be monitored by detecting de novo deleterious mutations in the genome of bulls used for artificial insemination. These results demonstrate the attractiveness of cattle as a model species in the post genomic era, particularly to confirm the genetic aetiology of isolated clinical case reports in humans.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Simmons D. The Use of Animal Models in Studying Genetic Disease: Transgenesis and Induced Mutation. Nature Education. 2008;1:70.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
