

Target of rapamycin signaling orchestrates growth-defense trade-offs in plants
- PMID: 28905991
- PMCID: PMC5711548
- DOI: 10.1111/nph.14785
Target of rapamycin signaling orchestrates growth-defense trade-offs in plants
Abstract
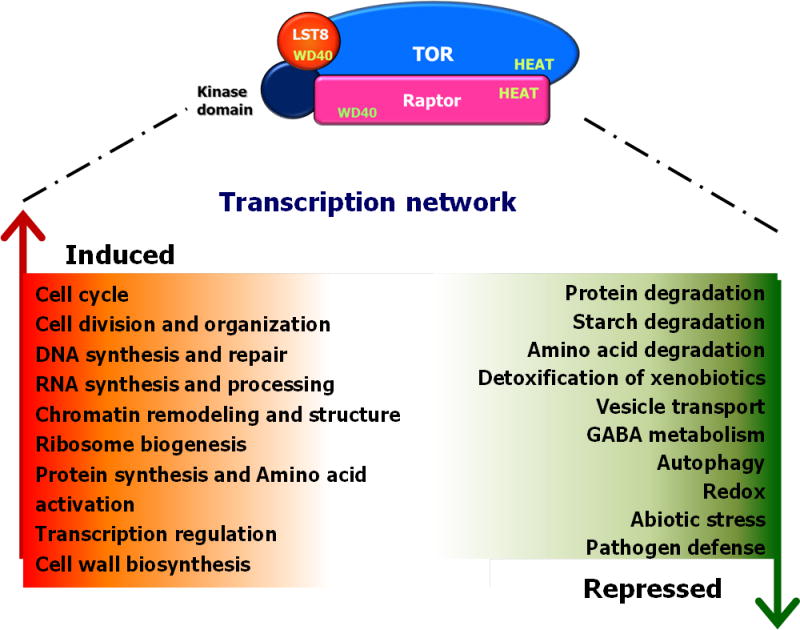
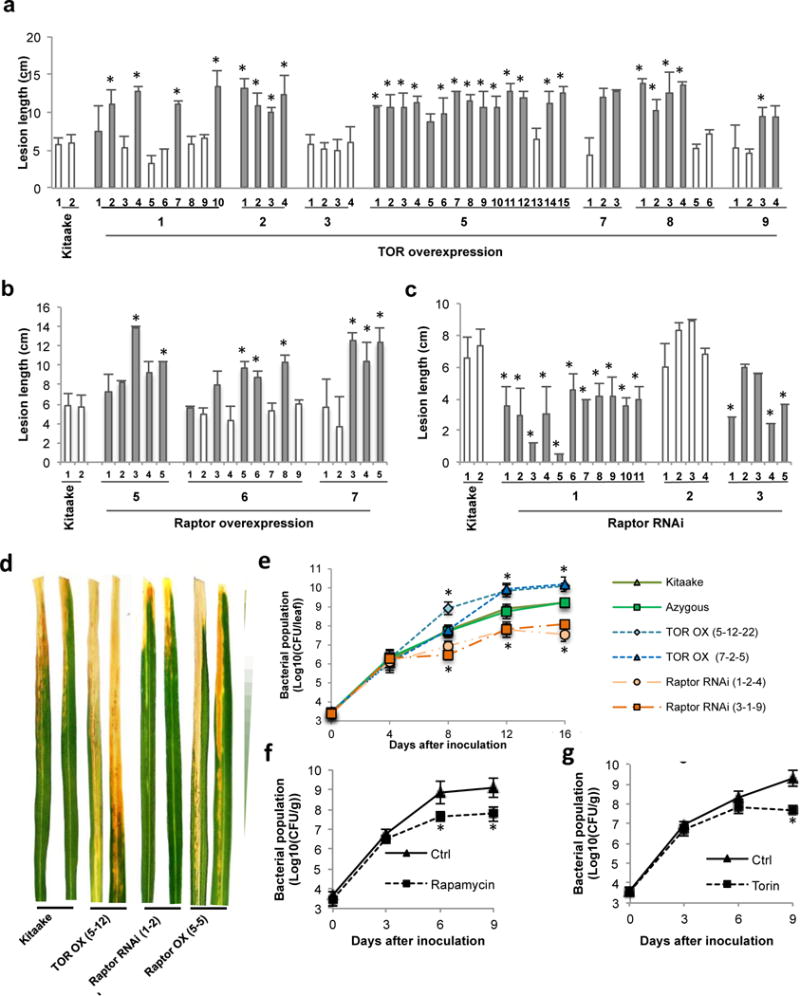
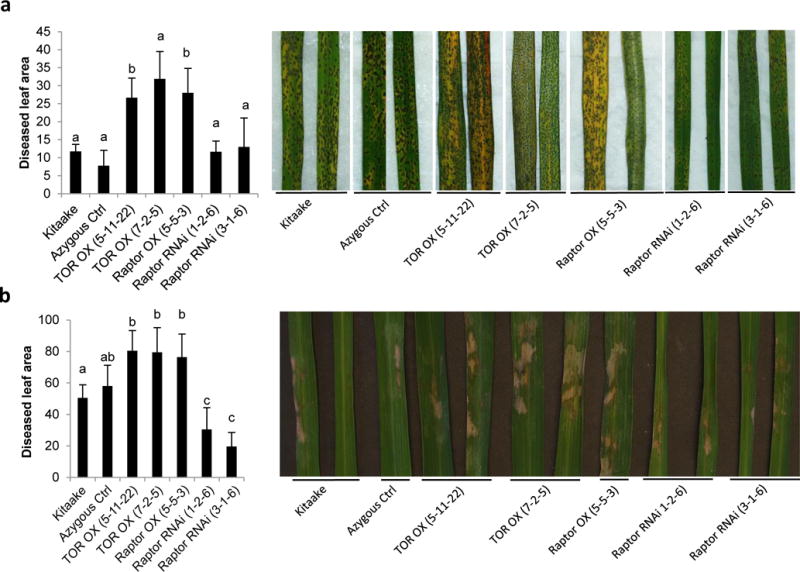
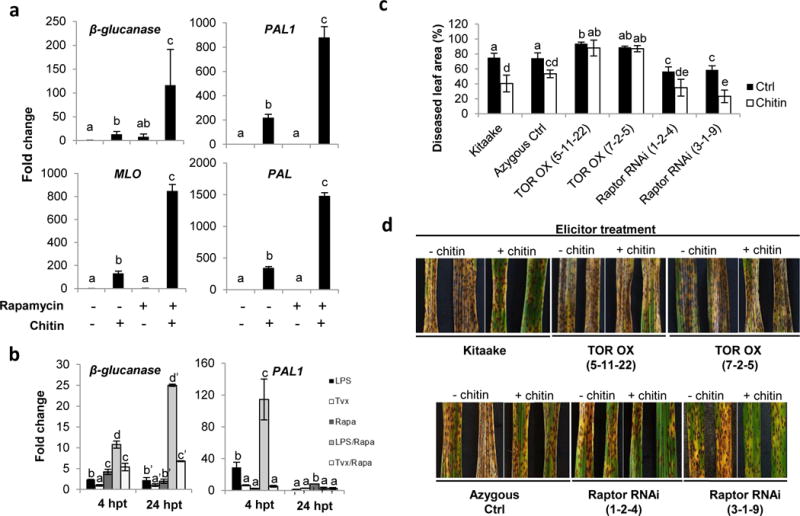
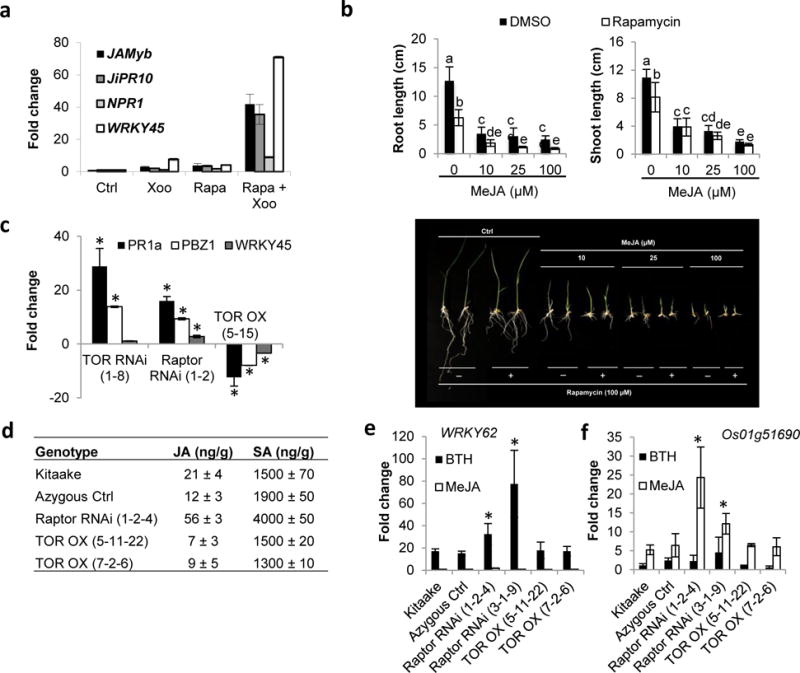
Plant defense to microbial pathogens is often accompanied by significant growth inhibition. How plants merge immune system function with normal growth and development is still poorly understood. Here, we investigated the role of target of rapamycin (TOR), an evolutionary conserved serine/threonine kinase, in the plant defense response. We used rice as a model system and applied a combination of chemical, genetic, genomic and cell-based analyses. We demonstrate that ectopic expression of TOR and Raptor (regulatory-associated protein of mTOR), a protein previously demonstrated to interact with TOR in Arabidopsis, positively regulates growth and development in rice. Transcriptome analysis of rice cells treated with the TOR-specific inhibitor rapamycin revealed that TOR not only dictates transcriptional reprogramming of extensive gene sets involved in central and secondary metabolism, cell cycle and transcription, but also suppresses many defense-related genes. TOR overexpression lines displayed increased susceptibility to both bacterial and fungal pathogens, whereas plants with reduced TOR signaling displayed enhanced resistance. Finally, we found that TOR antagonizes the action of the classic defense hormones salicylic acid and jasmonic acid. Together, these results indicate that TOR acts as a molecular switch for the activation of cell proliferation and plant growth at the expense of cellular immunity.
Keywords: growth-defense trade-offs; jasmonic acid; plant defense; rice (Oryza sativa); salicylic acid; target of rapamycin (TOR).
© 2017 The Authors. New Phytologist © 2017 New Phytologist Trust.
Figures

References
-
- Bjornson M, Dandekar AM, Chory J, Dehesh K. Brassinosteroid’s multi-modular interaction with the general stress network customizes stimulus-specific responses in Arabidopsis. Plant Science. 2016;250:165–177. - PubMed
-
- Brauc S, De Vooght E, Claeys M, Hofte M, Angenon G. Influence of over-expression of cytosolic aspartate aminotransferase on amino acid metabolism and defence responses against Botrytis cinerea infection in Arabidopsis thaliana. Journal of Plant Physiology. 2011;168(15):1813–1819. - PubMed
-
- Caldana C, Li Y, Leisse A, Zhang Y, Bartholomaeus L, Fernie AR, Willmitzer L, Giavalisco P. Systemic analysis of inducible target of rapamycin mutants reveal a general metabolic switch controlling growth in Arabidopsis thaliana. Plant Journal. 2013;73(6):897–909. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
