

Identification of a mouse Lactobacillus johnsonii strain with deconjugase activity against the FXR antagonist T-β-MCA
- PMID: 28910295
- PMCID: PMC5598929
- DOI: 10.1371/journal.pone.0183564
Identification of a mouse Lactobacillus johnsonii strain with deconjugase activity against the FXR antagonist T-β-MCA
Abstract
Bile salt hydrolase (BSH) activity against the bile acid tauro-beta-muricholic acid (T-β-MCA) was recently reported to mediate host bile acid, glucose, and lipid homeostasis via the farnesoid X receptor (FXR) signaling pathway. An earlier study correlated decreased Lactobacillus abundance in the cecum with increased concentrations of intestinal T-β-MCA, an FXR antagonist. While several studies have characterized BSHs in lactobacilli, deconjugation of T-β-MCA remains poorly characterized among members of this genus, and therefore it was unclear what strain(s) were responsible for this activity. Here, a strain of L. johnsonii with robust BSH activity against T-β-MCA in vitro was isolated from the cecum of a C57BL/6J mouse. A screening assay performed on a collection of 14 Lactobacillus strains from nine different species identified BSH substrate specificity for T-β-MCA only in two of three L. johnsonii strains. Genomic analysis of the two strains with this BSH activity revealed the presence of three bsh genes that are homologous to bsh genes in the previously sequenced human-associated strain L. johnsonii NCC533. Heterologous expression of several bsh genes in E. coli followed by enzymatic assays revealed broad differences in substrate specificity even among closely related bsh homologs, and suggests that the phylogeny of these enzymes does not closely correlate with substrate specificity. Predictive modeling allowed us to propose a potential mechanism driving differences in BSH activity for T-β-MCA in these homologs. Our data suggests that L. johnsonii regulates T-β-MCA levels in the mouse intestinal environment, and that this species may play a central role in FXR signaling in the mouse.
Conflict of interest statement
Figures
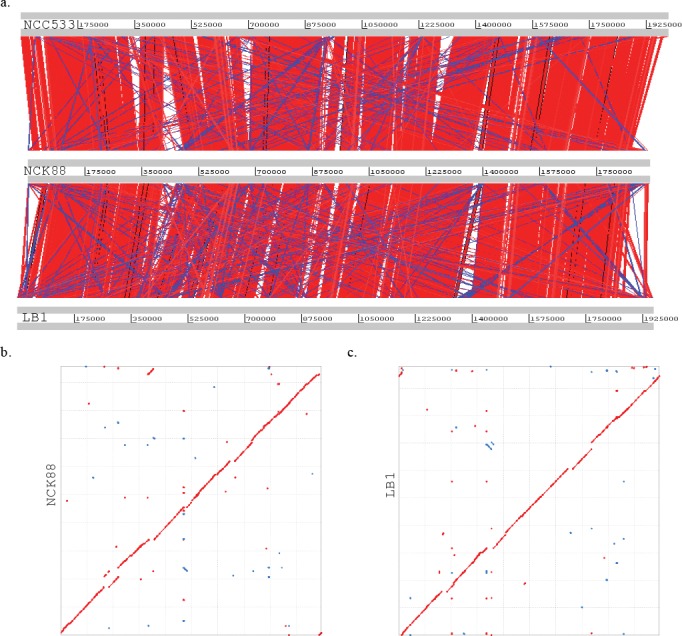
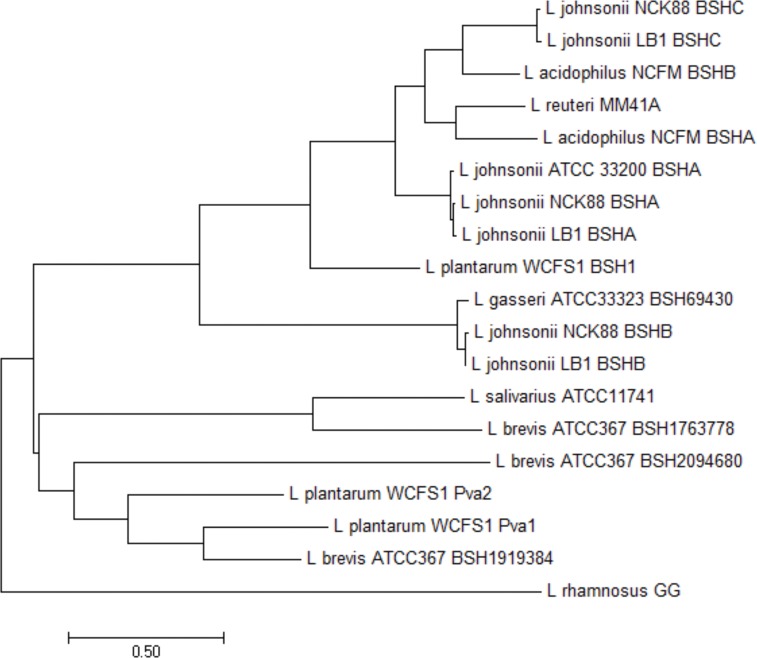
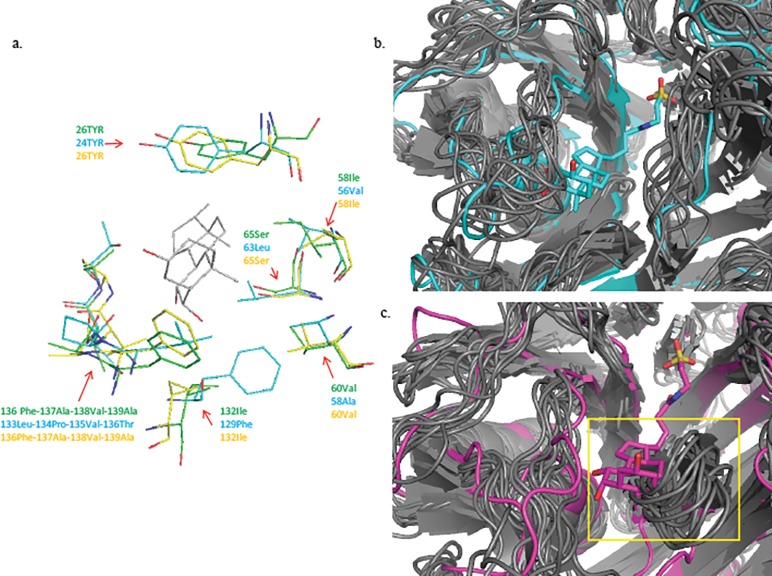
Similar articles
-
Cholesterol-lowering effect of bile salt hydrolase from a Lactobacillus johnsonii strain mediated by FXR pathway regulation.Food Funct. 2022 Jan 24;13(2):725-736. doi: 10.1039/d1fo03143k. Food Funct. 2022. PMID: 34935837
-
Molecular cloning, characterization and comparison of bile salt hydrolases from Lactobacillus johnsonii PF01.J Appl Microbiol. 2013 Jan;114(1):121-33. doi: 10.1111/jam.12027. Epub 2012 Oct 29. J Appl Microbiol. 2013. PMID: 23035872
-
Molecular cloning and characterization of a bile salt hydrolase from Lactobacillus acidophilus PF01.J Microbiol Biotechnol. 2008 Mar;18(3):449-56. J Microbiol Biotechnol. 2008. PMID: 18388461
-
An Intestinal Microbiota-Farnesoid X Receptor Axis Modulates Metabolic Disease.Gastroenterology. 2016 Nov;151(5):845-859. doi: 10.1053/j.gastro.2016.08.057. Epub 2016 Sep 14. Gastroenterology. 2016. PMID: 27639801 Free PMC article. Review.
-
Molecular features of bile salt hydrolases and relevance in human health.Biochim Biophys Acta Gen Subj. 2017 Jan;1861(1 Pt A):2981-2991. doi: 10.1016/j.bbagen.2016.09.024. Epub 2016 Sep 25. Biochim Biophys Acta Gen Subj. 2017. PMID: 27681686 Review.
Cited by
-
Galacto-Oligosaccharides Modulate the Juvenile Gut Microbiome and Innate Immunity To Improve Broiler Chicken Performance.mSystems. 2020 Jan 14;5(1):e00827-19. doi: 10.1128/mSystems.00827-19. mSystems. 2020. PMID: 31937680 Free PMC article.
-
Lactobacillus bile salt hydrolase substrate specificity governs bacterial fitness and host colonization.Proc Natl Acad Sci U S A. 2021 Feb 9;118(6):e2017709118. doi: 10.1073/pnas.2017709118. Proc Natl Acad Sci U S A. 2021. PMID: 33526676 Free PMC article.
-
Functional and Phylogenetic Diversity of BSH and PVA Enzymes.Microorganisms. 2021 Mar 31;9(4):732. doi: 10.3390/microorganisms9040732. Microorganisms. 2021. PMID: 33807488 Free PMC article. Review.
-
Evolutionary Analysis of Bile Acid-Conjugating Enzymes Reveals a Complex Duplication and Reciprocal Loss History.Genome Biol Evol. 2019 Nov 1;11(11):3256-3268. doi: 10.1093/gbe/evz238. Genome Biol Evol. 2019. PMID: 31670760 Free PMC article.
-
Effect of different bile acids on the intestine through enterohepatic circulation based on FXR.Gut Microbes. 2021 Jan-Dec;13(1):1949095. doi: 10.1080/19490976.2021.1949095. Gut Microbes. 2021. PMID: 34313539 Free PMC article. Review.
References
-
- Young VB. The role of the microbiome in human health and disease: an introduction for clinicians. BMJ. 2017;356: j831 doi: 10.1136/bmj.j831 - DOI - PubMed
-
- Geng W, Lin J. Bacterial bile salt hydrolase: an intestinal microbiome target for enhanced animal health. Anim Heal Res Rev. 2017/02/03. Cambridge University Press; 2016;17: 148–158. doi: 10.1017/S1466252316000153 - DOI - PubMed
-
- Ley RE, Bäckhed F, Turnbaugh P, Lozupone C a, Knight RD, Gordon JI. Obesity alters gut microbial ecology. Proc Natl Acad Sci U S A. 2005;102: 11070–5. doi: 10.1073/pnas.0504978102 - DOI - PMC - PubMed
-
- Ley R, Turnbaugh P, Klein S, Gordon J. Microbial ecology: human gut microbes associated with obesity. Nature. 2006;444: 1022–1023. doi: 10.1038/4441022a - DOI - PubMed
-
- Turnbaugh PJ, Ley RE, Mahowald M a, Magrini V, Mardis ER, Gordon JI. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature. 2006;444: 1027–31. doi: 10.1038/nature05414 - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
