

Aminoacyl-tRNA quality control is required for efficient activation of the TOR pathway regulator Gln3p
- PMID: 28910581
- PMCID: PMC6103723
- DOI: 10.1080/15476286.2017.1379635
Aminoacyl-tRNA quality control is required for efficient activation of the TOR pathway regulator Gln3p
Abstract
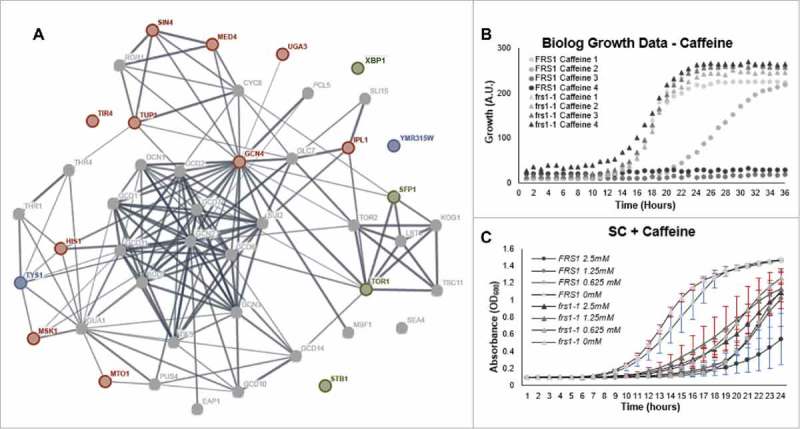
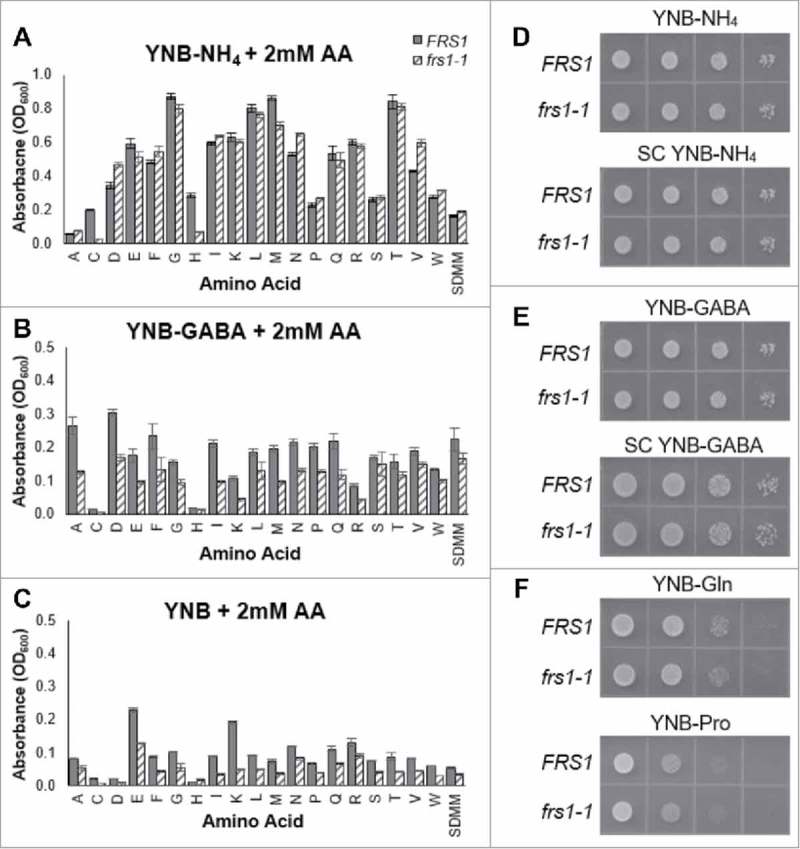
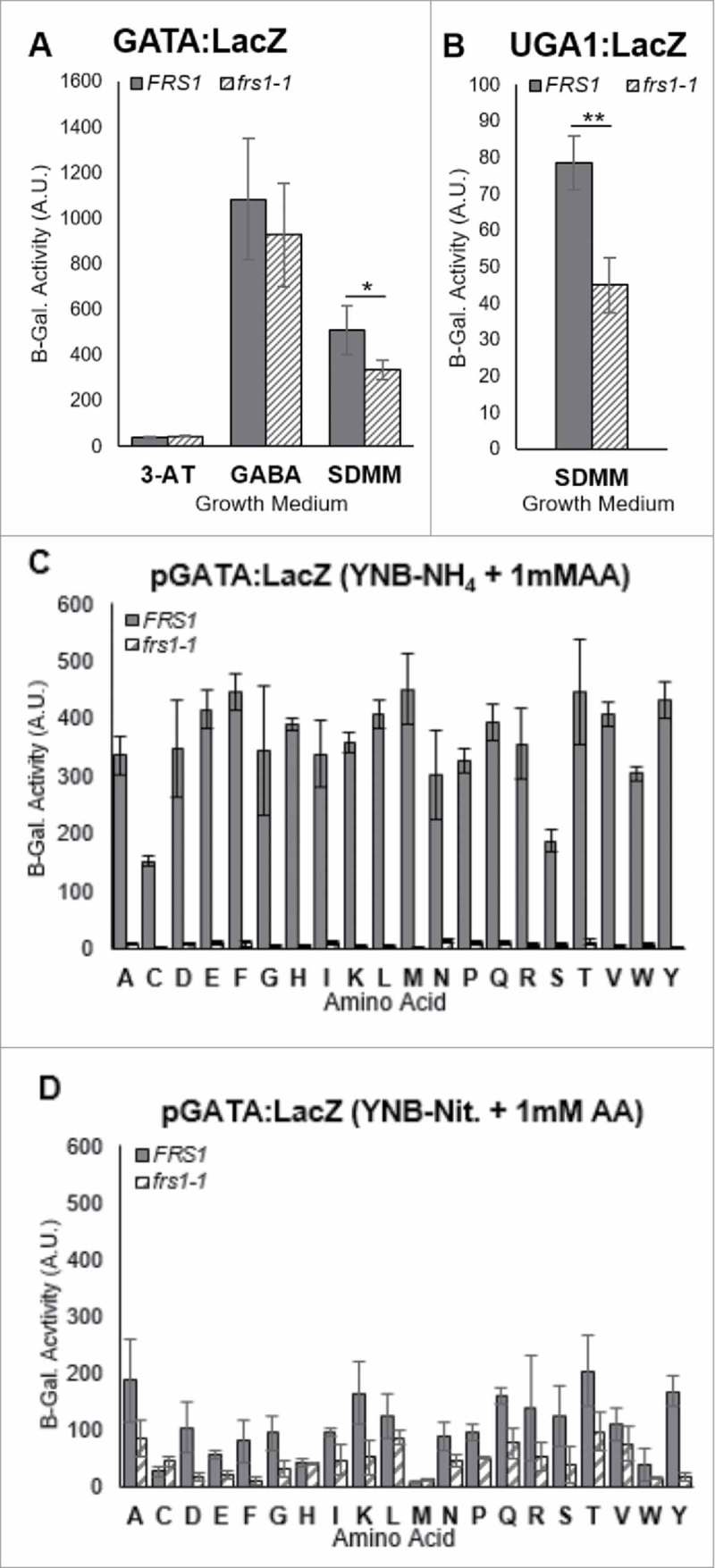
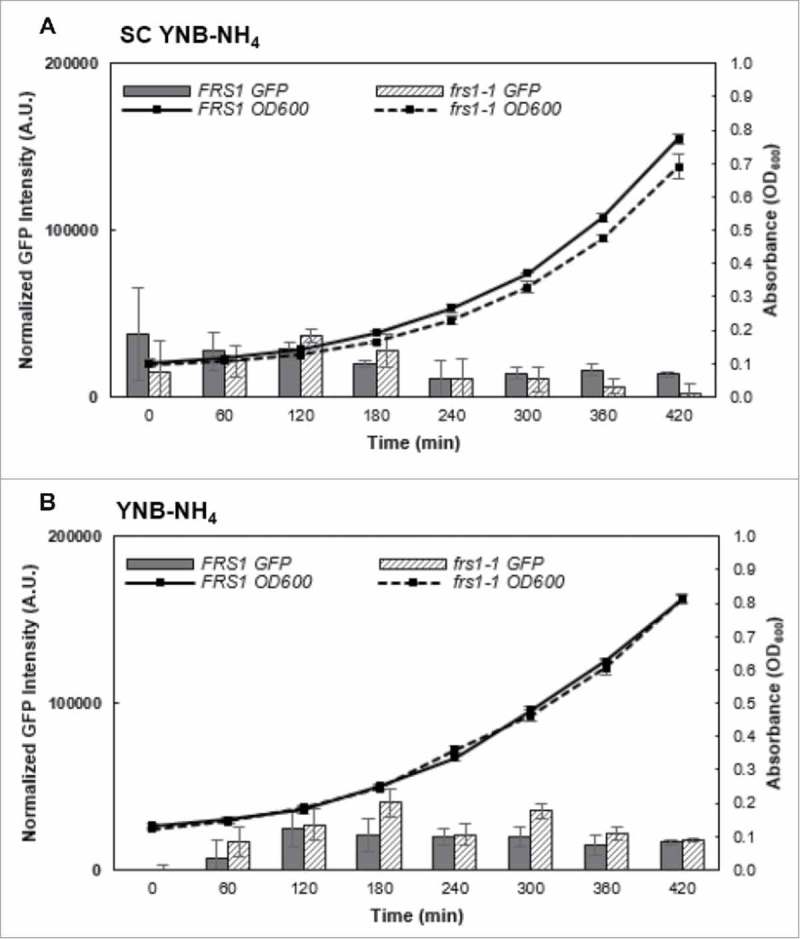
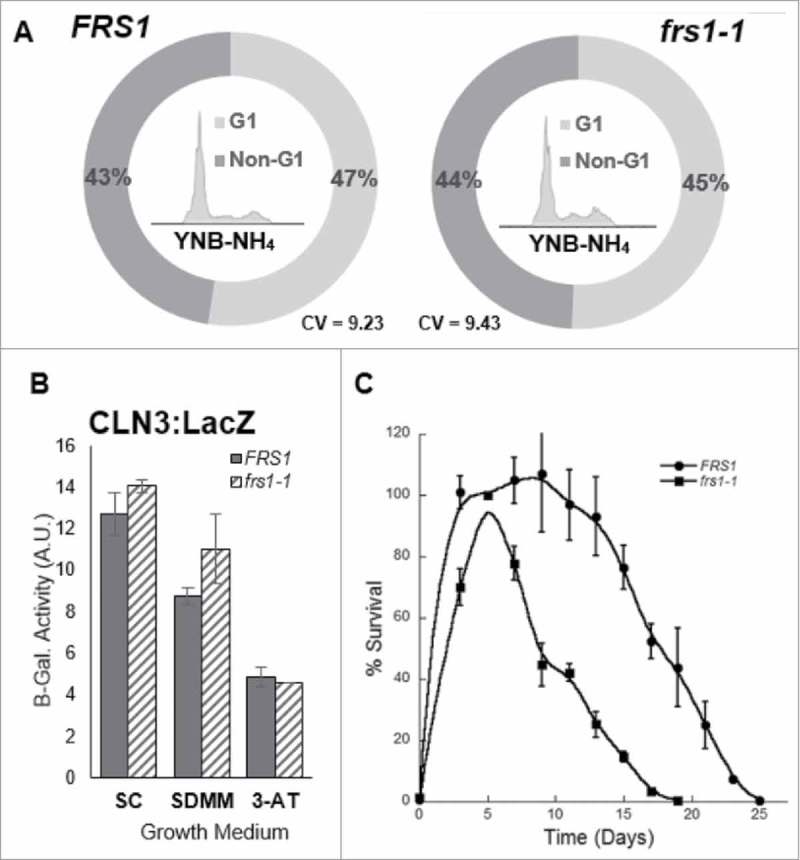
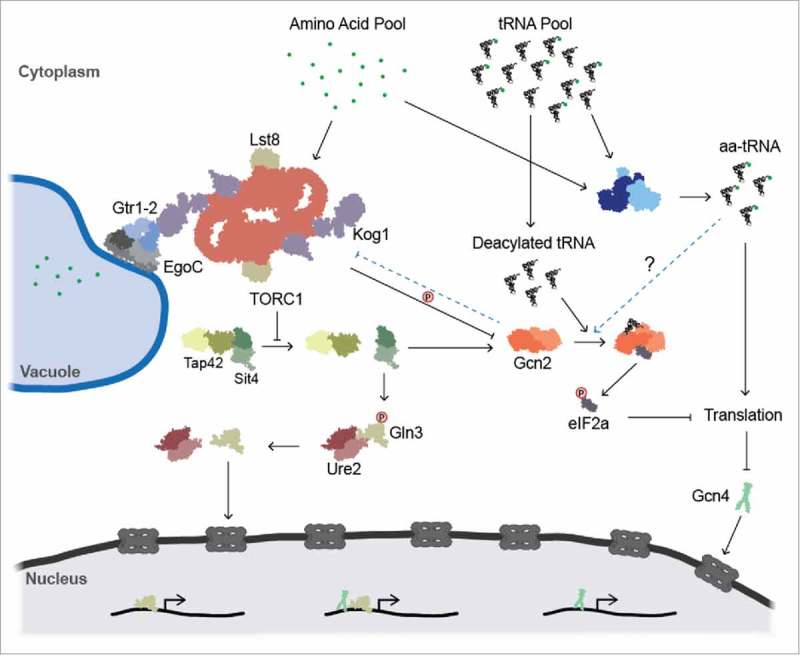
The aminoacylation status of the cellular tRNA pool regulates both general amino acid control (GAAC) and target of rapamycin (TOR) stress response pathways in yeast. Consequently, fidelity of translation at the level of aminoacyl-tRNA synthesis plays a central role in determining accuracy and sensitivity of stress responses. To investigate effects of translational quality control (QC) on cell physiology under stress conditions, phenotypic microarray analyses were used to identify changes in QC deficient cells. Nitrogen source growth assays showed QC deficient yeast grew differently compared to WT. The QC deficient strain was more tolerant to caffeine treatment than wild type through altered interactions with the TOR and GAAC pathways. Increased caffeine tolerance of the QC deficient strain was consistent with the observation that the activity of Gln3p, a transcription factor controlled by the TOR pathway, is decreased in the QC deficient strain compared to WT. GCN4 translation, which is typically repressed in the absence of nutritional stress, was enhanced in the QC deficient strain through TOR inhibition. QC did not impact cell cycle regulation; however, the chronological lifespan of QC deficient yeast strains decreased compared to wild type, likely due to translational errors and alteration of the TOR-associated regulon. These findings support the idea that changes in translational fidelity provide a mechanism of cellular adaptation by modulating TOR activity. This, in turn, supports a central role for aminoacyl-tRNA synthesis QC in the integrated stress response by maintaining the proper aa-tRNA pools necessary to coordinate the GAAC and TOR.
Keywords: Aminoacyl-tRNA; GAAC; TOR; nutritional stress; tRNA editing.
Figures
Similar articles
-
Integration of general amino acid control and target of rapamycin (TOR) regulatory pathways in nitrogen assimilation in yeast.J Biol Chem. 2010 May 28;285(22):16893-911. doi: 10.1074/jbc.M110.121947. Epub 2010 Mar 16. J Biol Chem. 2010. PMID: 20233714 Free PMC article.
-
A novel role for protein kinase Gcn2 in yeast tolerance to intracellular acid stress.Biochem J. 2012 Jan 1;441(1):255-64. doi: 10.1042/BJ20111264. Biochem J. 2012. PMID: 21919885
-
Translation quality control is critical for bacterial responses to amino acid stress.Proc Natl Acad Sci U S A. 2016 Feb 23;113(8):2252-7. doi: 10.1073/pnas.1525206113. Epub 2016 Feb 8. Proc Natl Acad Sci U S A. 2016. PMID: 26858451 Free PMC article.
-
Translational fidelity and mistranslation in the cellular response to stress.Nat Microbiol. 2017 Aug 24;2:17117. doi: 10.1038/nmicrobiol.2017.117. Nat Microbiol. 2017. PMID: 28836574 Free PMC article. Review.
-
Regulation of RNA polymerase III transcription by Maf1 protein.Acta Biochim Pol. 2008;55(2):215-25. Epub 2008 Jun 17. Acta Biochim Pol. 2008. PMID: 18560610 Review.
Cited by
-
Methionyl-tRNA synthetase synthetic and proofreading activities are determinants of antibiotic persistence.Front Microbiol. 2024 Mar 27;15:1384552. doi: 10.3389/fmicb.2024.1384552. eCollection 2024. Front Microbiol. 2024. PMID: 38601944 Free PMC article.
-
The leucine-NH4+ uptake regulator Any1 limits growth as part of a general amino acid control response to loss of La protein by fission yeast.PLoS One. 2021 Jun 21;16(6):e0253494. doi: 10.1371/journal.pone.0253494. eCollection 2021. PLoS One. 2021. PMID: 34153074 Free PMC article.
-
Yeast Chronological Lifespan: Longevity Regulatory Genes and Mechanisms.Cells. 2022 May 23;11(10):1714. doi: 10.3390/cells11101714. Cells. 2022. PMID: 35626750 Free PMC article. Review.
-
Saccharomyces cerevisiae and Caffeine Implications on the Eukaryotic Cell.Nutrients. 2020 Aug 13;12(8):2440. doi: 10.3390/nu12082440. Nutrients. 2020. PMID: 32823708 Free PMC article. Review.
-
How to Screen and Prevent Metabolic Syndrome in Patients of PCOS Early: Implications From Metabolomics.Front Endocrinol (Lausanne). 2021 Jun 2;12:659268. doi: 10.3389/fendo.2021.659268. eCollection 2021. Front Endocrinol (Lausanne). 2021. PMID: 34149613 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases