

Structural accommodation of ribonucleotide incorporation by the DNA repair enzyme polymerase Mu
- PMID: 28911097
- PMCID: PMC5587726
- DOI: 10.1093/nar/gkx527
Structural accommodation of ribonucleotide incorporation by the DNA repair enzyme polymerase Mu
Abstract
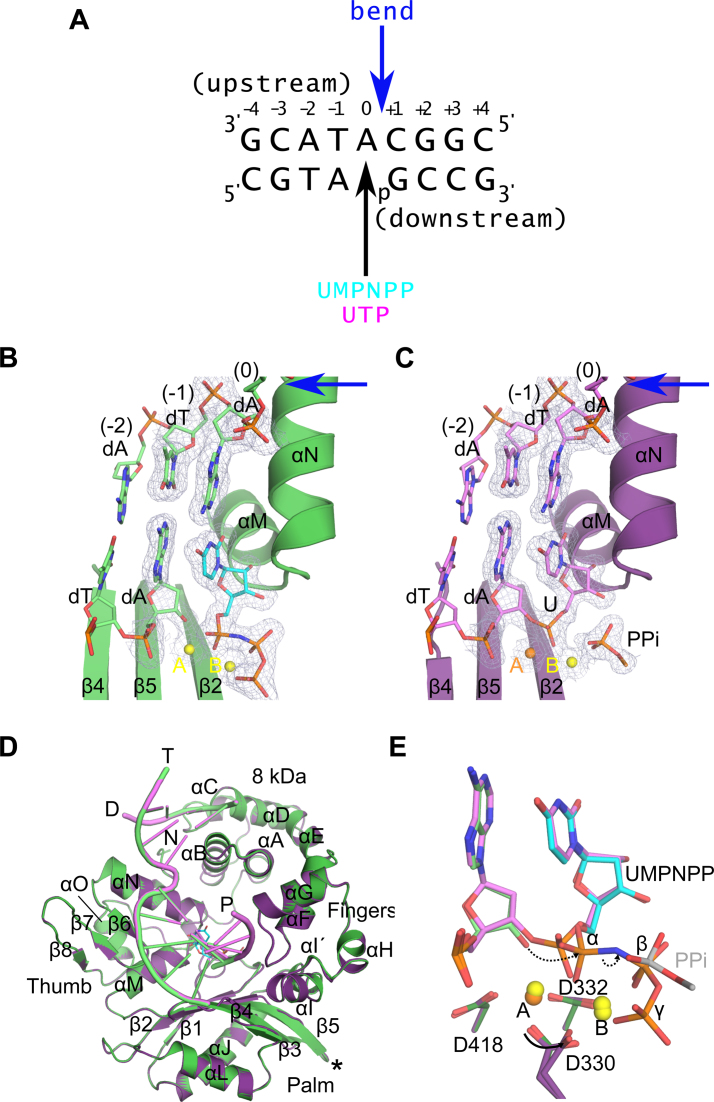
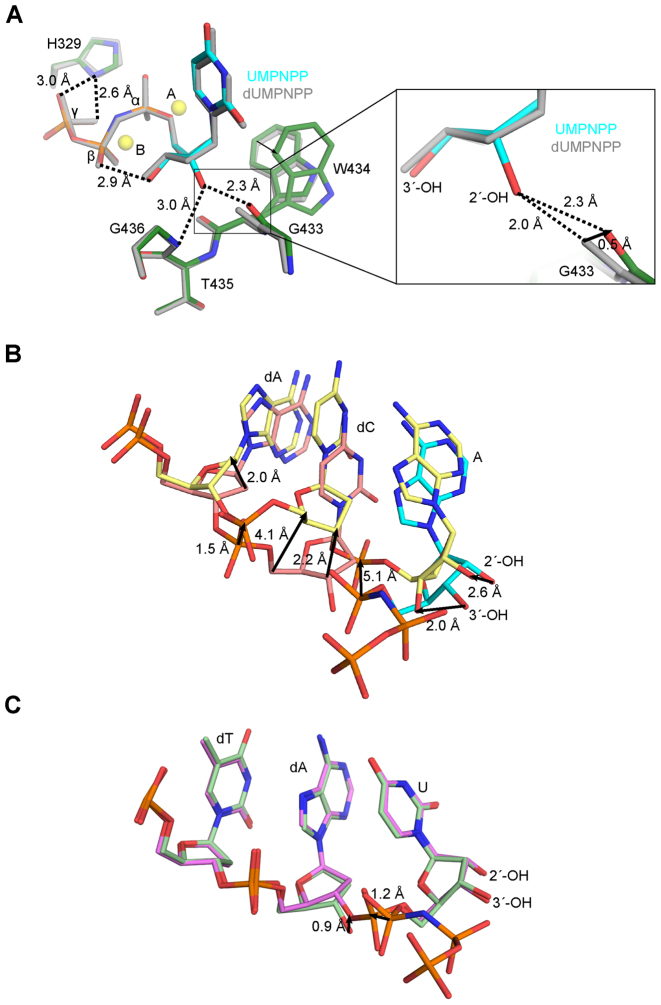
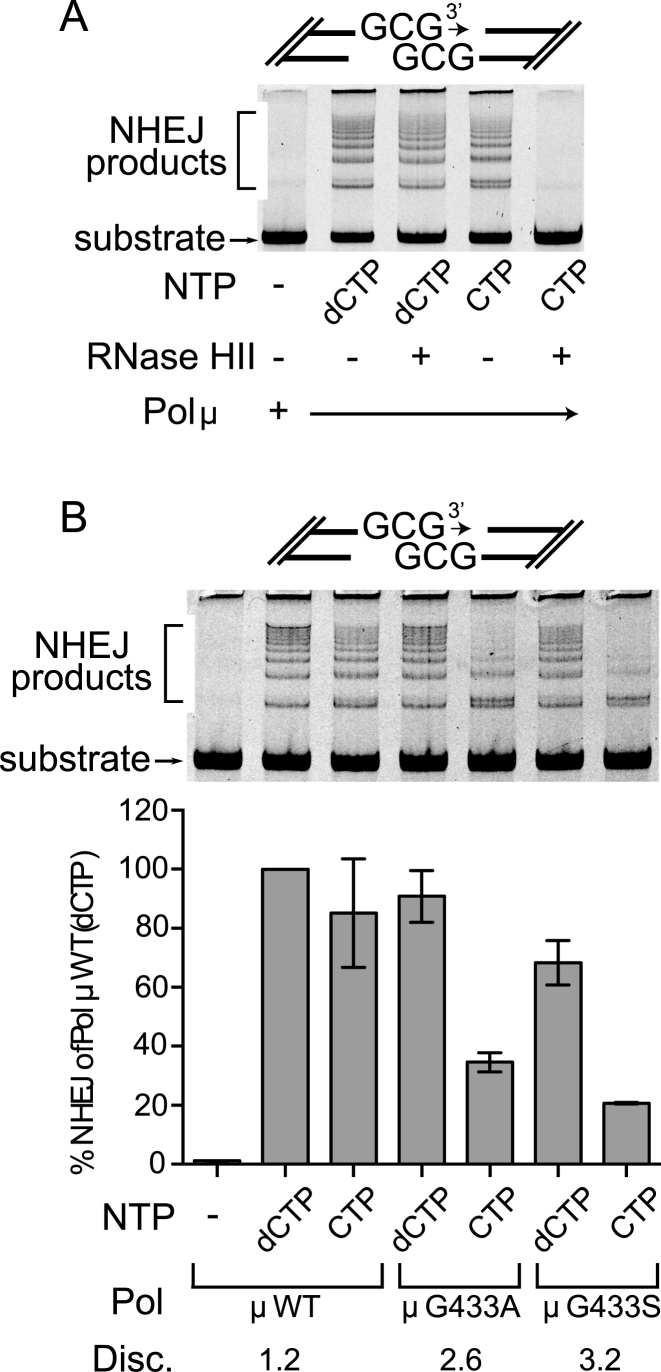
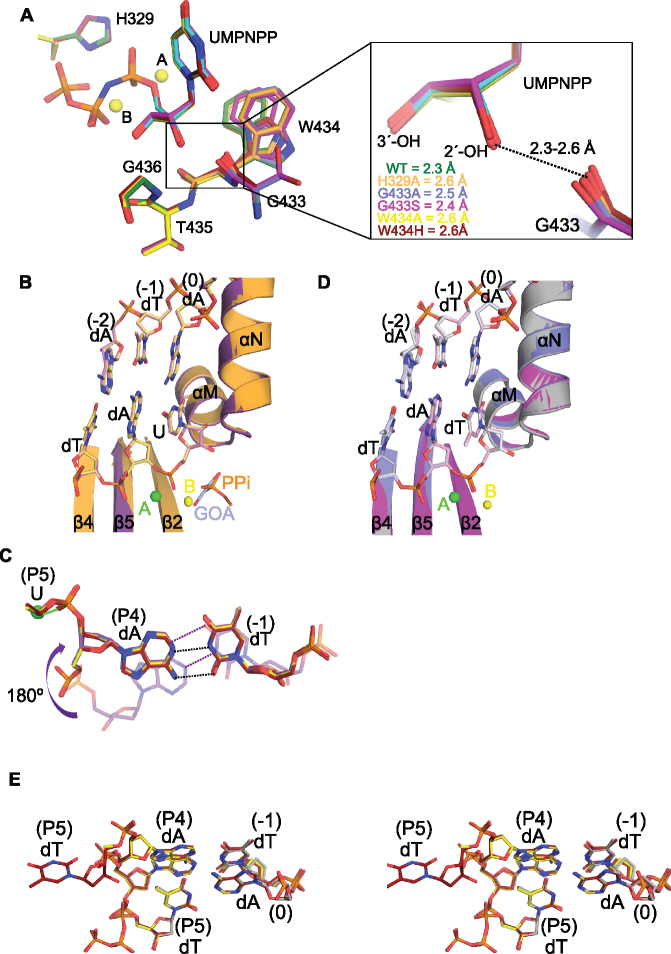
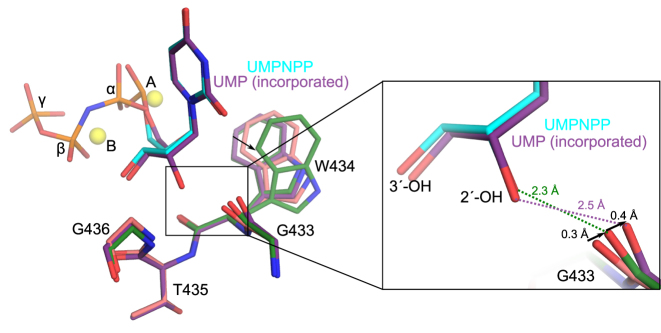
While most DNA polymerases discriminate against ribonucleotide triphosphate (rNTP) incorporation very effectively, the Family X member DNA polymerase μ (Pol μ) incorporates rNTPs almost as efficiently as deoxyribonucleotides. To gain insight into how this occurs, here we have used X-ray crystallography to describe the structures of pre- and post-catalytic complexes of Pol μ with a ribonucleotide bound at the active site. These structures reveal that Pol μ binds and incorporates a rNTP with normal active site geometry and no distortion of the DNA substrate or nucleotide. Moreover, a comparison of rNTP incorporation kinetics by wildtype and mutant Pol μ indicates that rNTP accommodation involves synergistic interactions with multiple active site residues not found in polymerases with greater discrimination. Together, the results are consistent with the hypothesis that rNTP incorporation by Pol μ is advantageous in gap-filling synthesis during DNA double strand break repair by nonhomologous end joining, particularly in nonreplicating cells containing very low deoxyribonucleotide concentrations.
Published by Oxford University Press on behalf of Nucleic Acids Research 2017.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
