

Genome-wide analysis of influenza viral RNA and nucleoprotein association
- PMID: 28911100
- PMCID: PMC5587783
- DOI: 10.1093/nar/gkx584
Genome-wide analysis of influenza viral RNA and nucleoprotein association
Abstract
Influenza A virus (IAV) genomes are composed of eight single-stranded RNA segments that are coated by viral nucleoprotein (NP) molecules. Classically, the interaction between NP and viral RNA (vRNA) is depicted as a uniform pattern of 'beads on a string'. Using high-throughput sequencing of RNA isolated by crosslinking immunoprecipitation (HITS-CLIP), we identified the vRNA binding profiles of NP for two H1N1 IAV strains in virions. Contrary to the prevailing model for vRNA packaging, NP does not bind vRNA uniformly in the A/WSN/1933 and A/California/07/2009 strains, but instead each vRNA segment exhibits a unique binding profile, containing sites that are enriched or poor in NP association. Intriguingly, both H1N1 strains have similar yet distinct NP binding profiles despite extensive sequence conservation. Peaks identified by HITS-CLIP were verified as true NP binding sites based on insensitivity to DNA antisense oligonucleotide-mediated RNase H digestion. Moreover, nucleotide content analysis of NP peaks revealed that these sites are relatively G-rich and U-poor compared to the genome-wide nucleotide content, indicating an as-yet unidentified sequence bias for NP association in vivo. Taken together, our genome-wide study of NP-vRNA interaction has implications for the understanding of influenza vRNA architecture and genome packaging.
© The Author(s) 2017. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
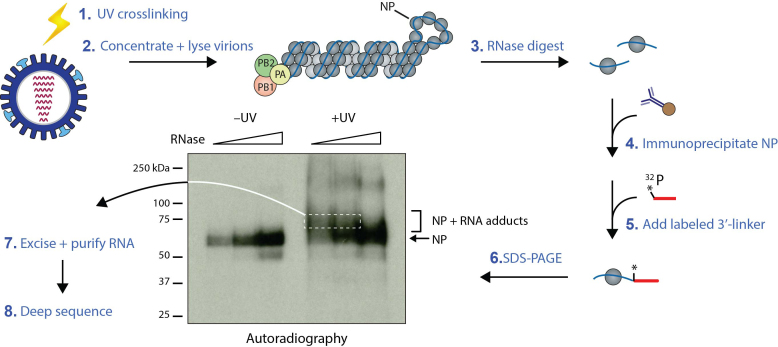

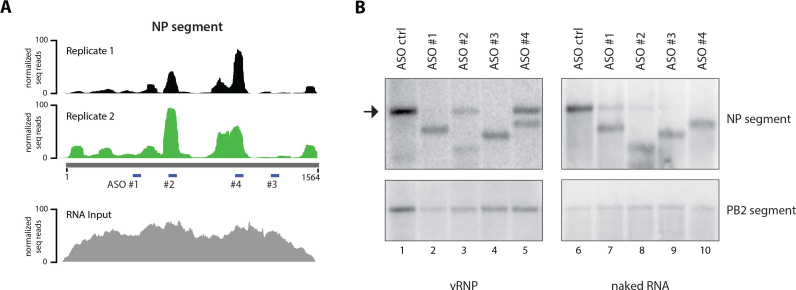

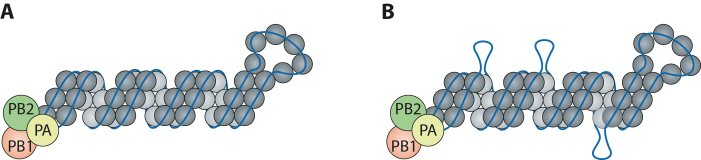
References
-
- Arranz R., Coloma R., Chichon F.J., Conesa J.J., Carrascosa J.L., Valpuesta J.M., Ortin J., Martin-Benito J.. The structure of native influenza virion ribonucleoproteins. Science. 2012; 338:1634–1637. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
