

The IRES5'UTR of the dicistrovirus cricket paralysis virus is a type III IRES containing an essential pseudoknot structure
- PMID: 28911115
- PMCID: PMC5587806
- DOI: 10.1093/nar/gkx622
The IRES5'UTR of the dicistrovirus cricket paralysis virus is a type III IRES containing an essential pseudoknot structure
Abstract
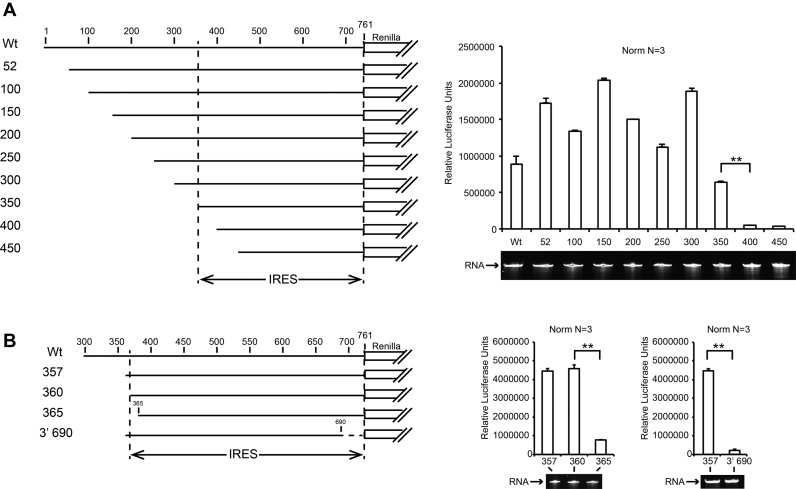
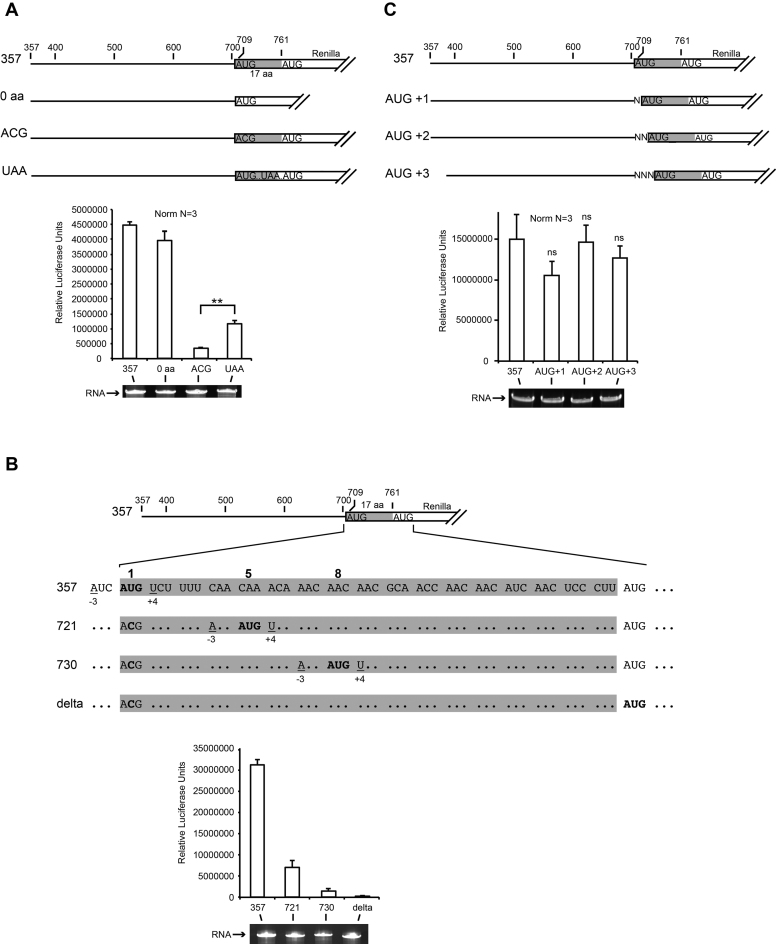
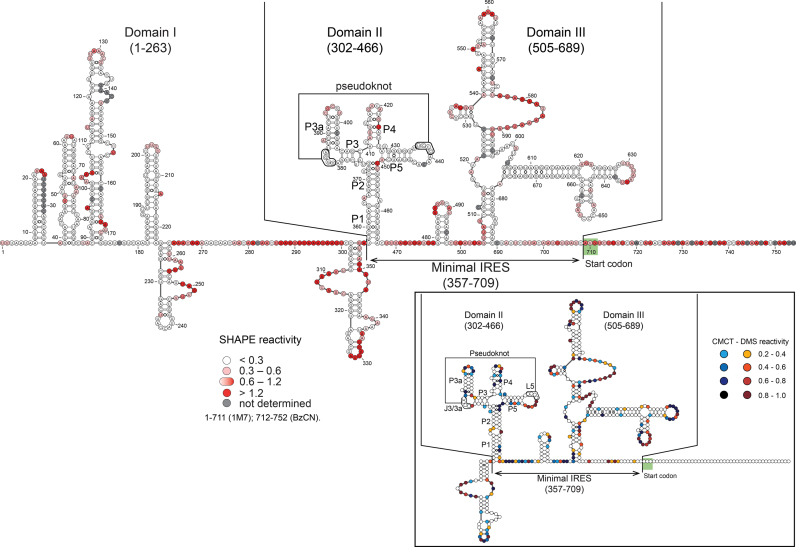
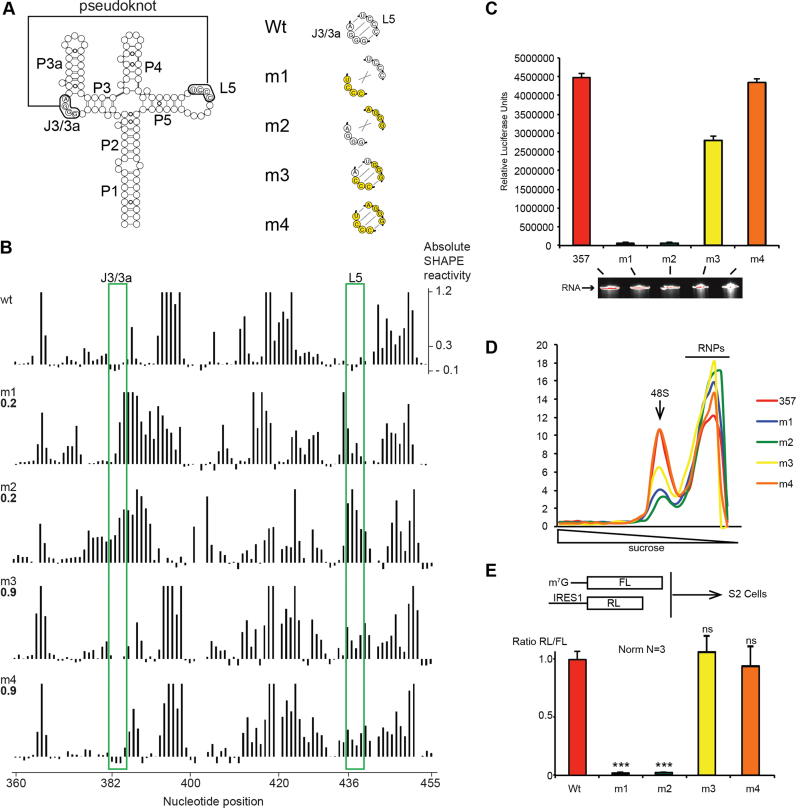
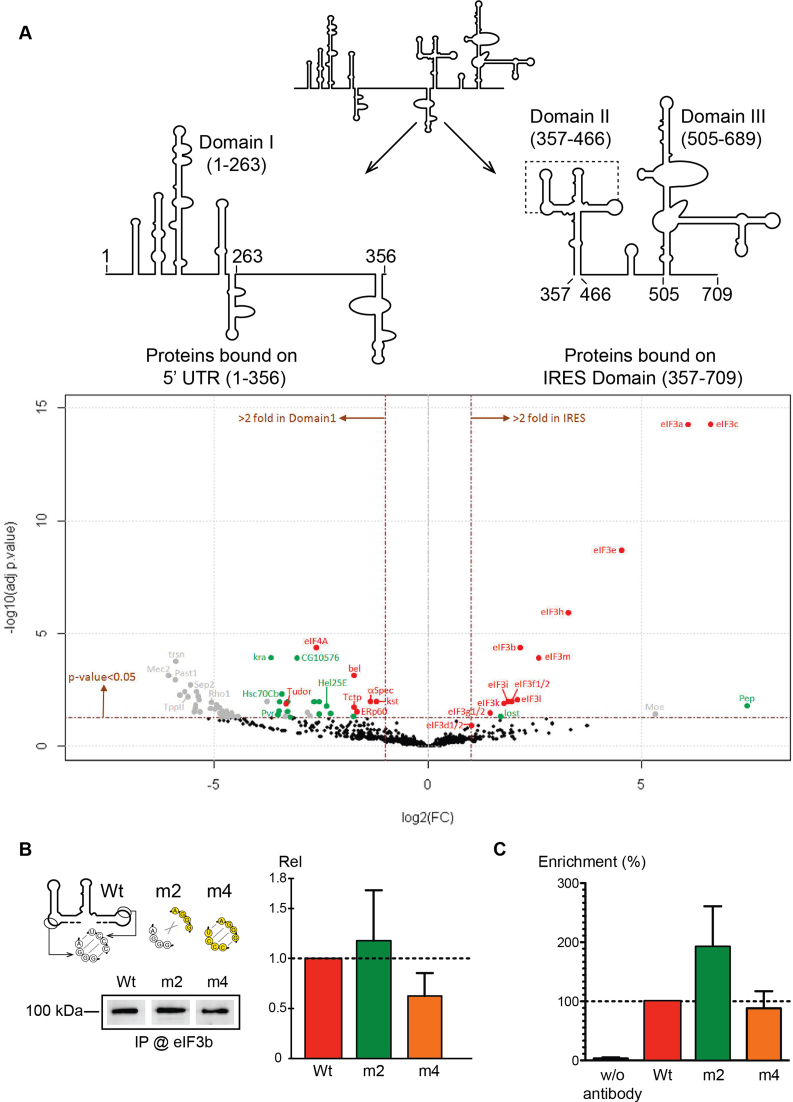
Cricket paralysis virus (CrPV) is a dicistrovirus. Its positive-sense single-stranded RNA genome contains two internal ribosomal entry sites (IRESs). The 5' untranslated region (5'UTR) IRES5'UTR mediates translation of non-structural proteins encoded by ORF1 whereas the well-known intergenic region (IGR) IRESIGR is required for translation of structural proteins from open reading frame 2 in the late phase of infection. Concerted action of both IRES is essential for host translation shut-off and viral translation. IRESIGR has been extensively studied, in contrast the IRES5'UTR remains largely unexplored. Here, we define the minimal IRES element required for efficient translation initiation in drosophila S2 cell-free extracts. We show that IRES5'UTR promotes direct recruitment of the ribosome on the cognate viral AUG start codon without any scanning step, using a Hepatitis-C virus-related translation initiation mechanism. Mass spectrometry analysis revealed that IRES5'UTR recruits eukaryotic initiation factor 3, confirming that it belongs to type III class of IRES elements. Using Selective 2'-hydroxyl acylation analyzed by primer extension and DMS probing, we established a secondary structure model of 5'UTR and of the minimal IRES5'UTR. The IRES5'UTR contains a pseudoknot structure that is essential for proper folding and ribosome recruitment. Overall, our results pave the way for studies addressing the synergy and interplay between the two IRES from CrPV.
© The Author(s) 2017. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Etchison D., Milburn S.C., Edery I., Sonenberg N., Hershey J.W.. Inhibition of HeLa cell protein synthesis following poliovirus infection correlates with the proteolysis of a 220, 000-dalton polypeptide associated with eucaryotic initiation factor 3 and a cap binding protein complex. J. Biol. Chem. 1982; 257:14806–14810. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
