

Senescence is a Spi1-induced anti-proliferative mechanism in primary hematopoietic cells
- PMID: 28912174
- PMCID: PMC5664389
- DOI: 10.3324/haematol.2016.157636
Senescence is a Spi1-induced anti-proliferative mechanism in primary hematopoietic cells
Abstract
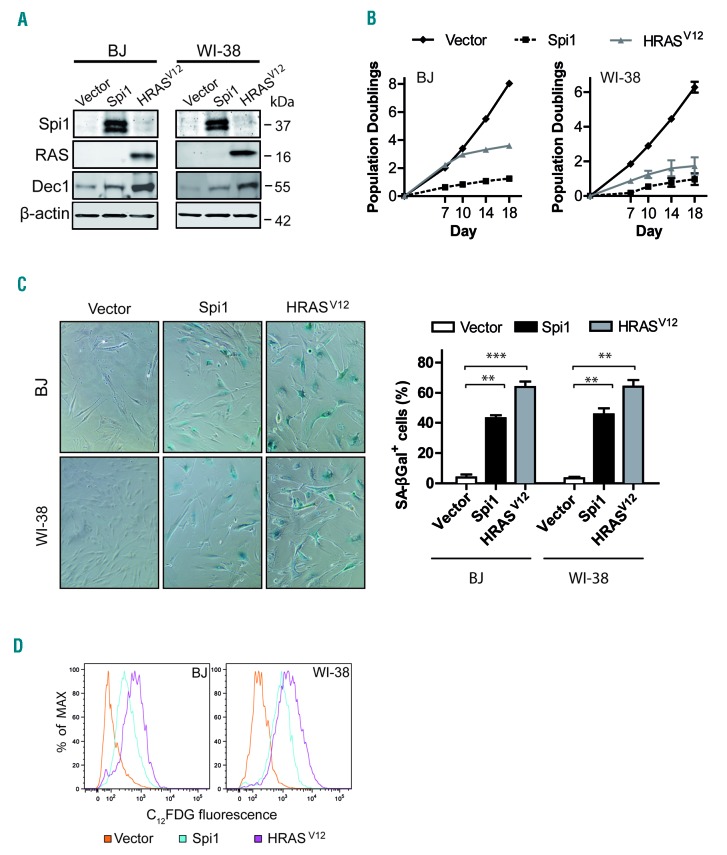
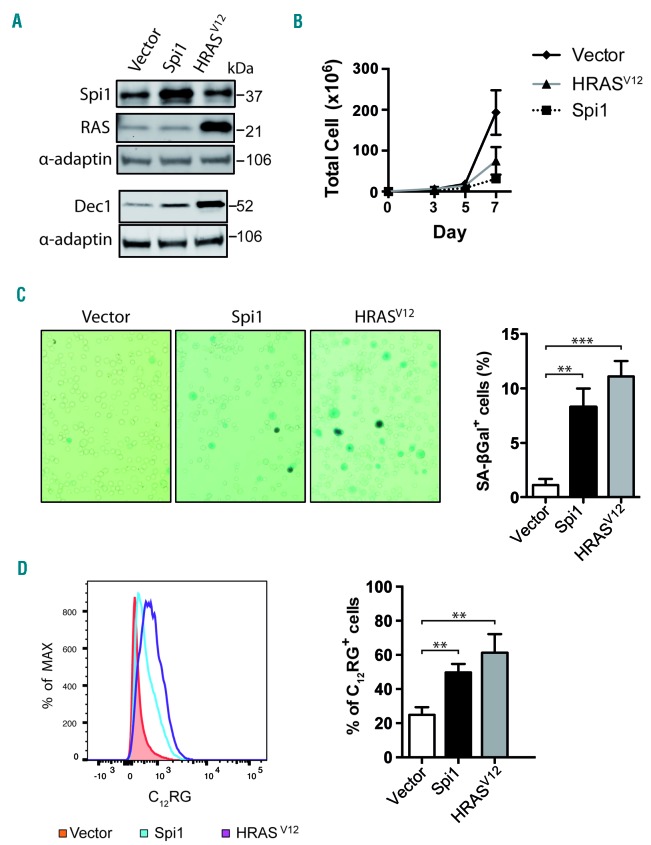
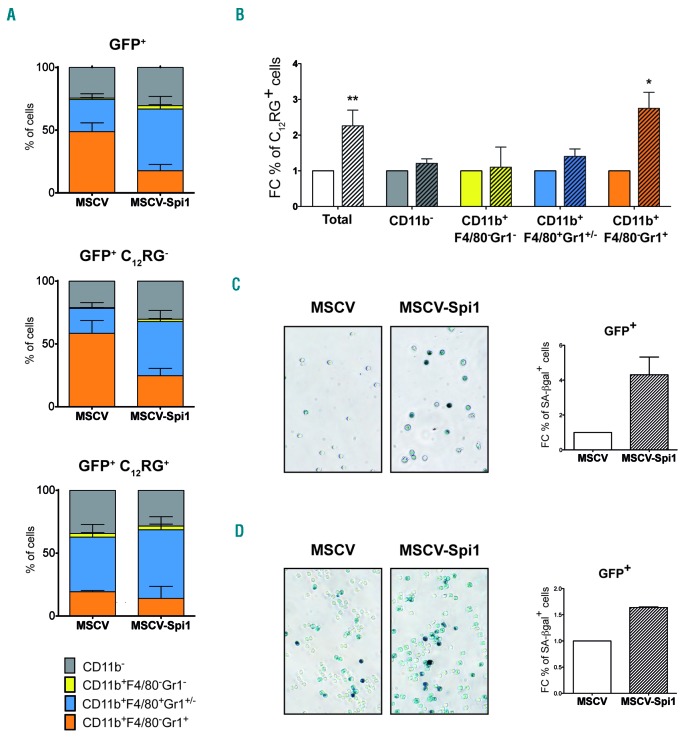
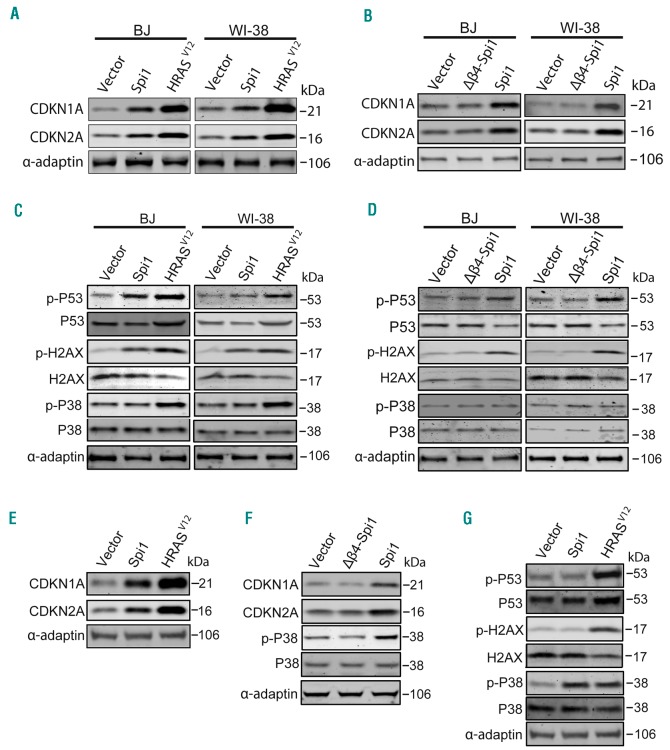
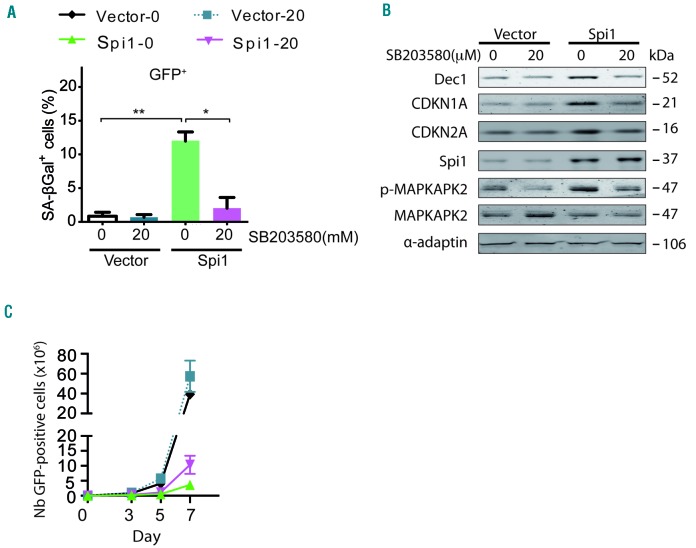
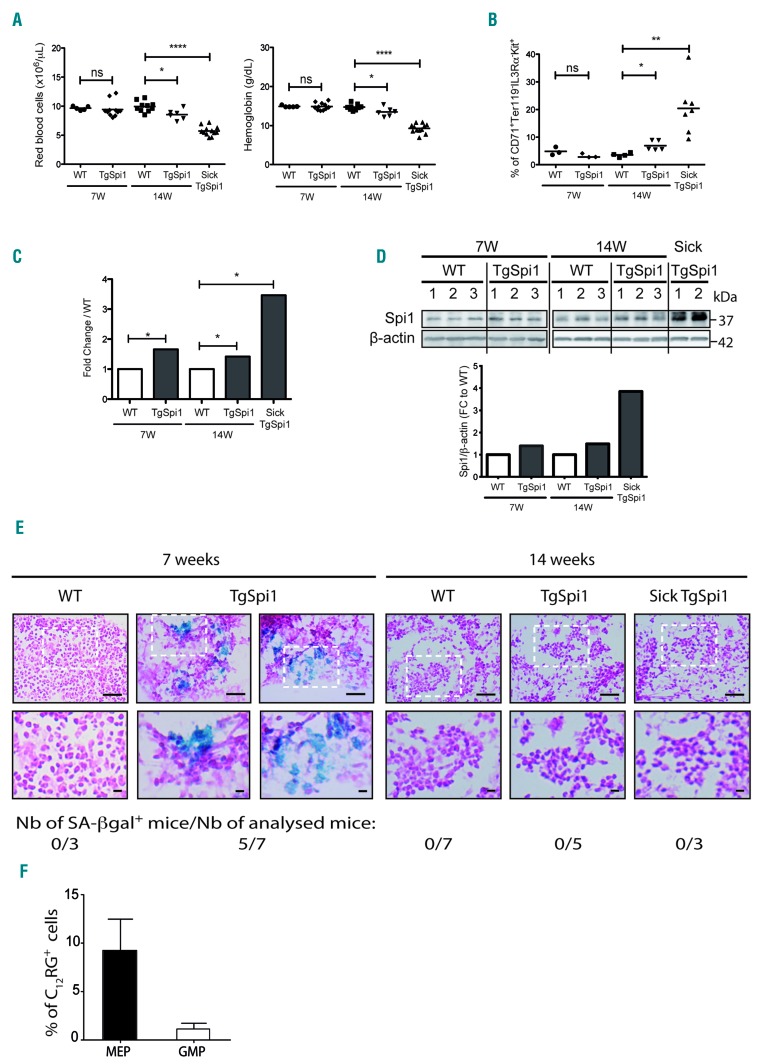
Transcriptional deregulation caused by epigenetic or genetic alterations is a major cause of leukemic transformation. The Spi1/PU.1 transcription factor is a key regulator of many steps of hematopoiesis, and limits self-renewal of hematopoietic stem cells. The deregulation of its expression or activity contributes to leukemia, in which Spi1 can be either an oncogene or a tumor suppressor. Herein we explored whether cellular senescence, an anti-tumoral pathway that restrains cell proliferation, is a mechanism by which Spi1 limits hematopoietic cell expansion, and thus prevents the development of leukemia. We show that Spi1 overexpression triggers cellular senescence both in primary fibroblasts and hematopoietic cells. Erythroid and myeloid lineages are both prone to Spi1-induced senescence. In hematopoietic cells, Spi1-induced senescence requires its DNA-binding activity and a functional p38MAPK14 pathway but is independent of a DNA-damage response. In contrast, in fibroblasts, Spi1-induced senescence is triggered by a DNA-damage response. Importantly, using our well-established Spi1 transgenic leukemia mouse model, we demonstrate that Spi1 overexpression also induces senescence in erythroid progenitors of the bone marrow in vivo before the onset of the pre-leukemic phase of erythroleukemia. Remarkably, the senescence response is lost during the progression of the disease and erythroid blasts do not display a higher expression of Dec1 and CDKN1A, two of the induced senescence markers in young animals. These results bring indirect evidence that leukemia develops from cells which have bypassed Spi1-induced senescence. Overall, our results reveal senescence as a Spi1-induced anti-proliferative mechanism that may be a safeguard against the development of acute myeloid leukemia.
Copyright© Ferrata Storti Foundation.
Figures
References
-
- Kim HG, de Guzman CG, Swindle CS, et al. The ETS family transcription factor PU.1 is necessary for the maintenance of fetal liver hematopoietic stem cells. Blood. 2004;104(13):3894–3900. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
