

IF2 and unique features of initiator tRNAfMet help establish the translational reading frame
- PMID: 28914580
- PMCID: PMC6103701
- DOI: 10.1080/15476286.2017.1379636
IF2 and unique features of initiator tRNAfMet help establish the translational reading frame
Abstract
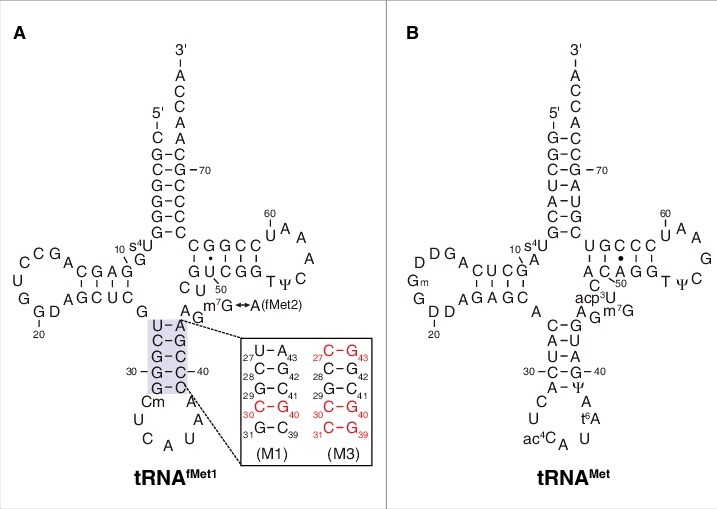
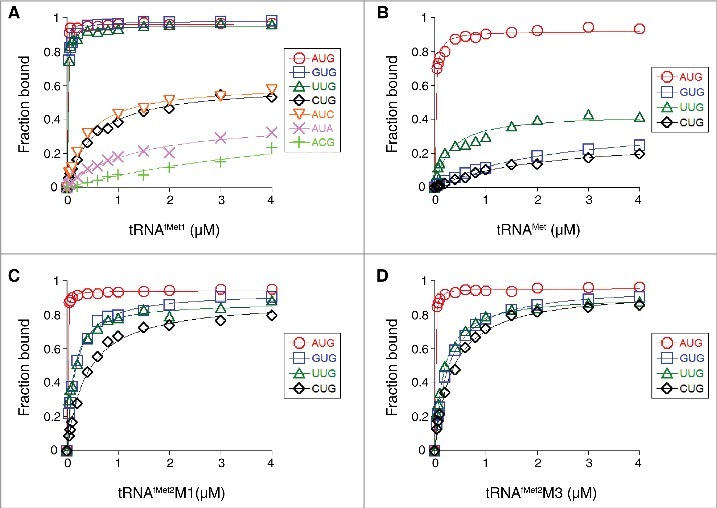
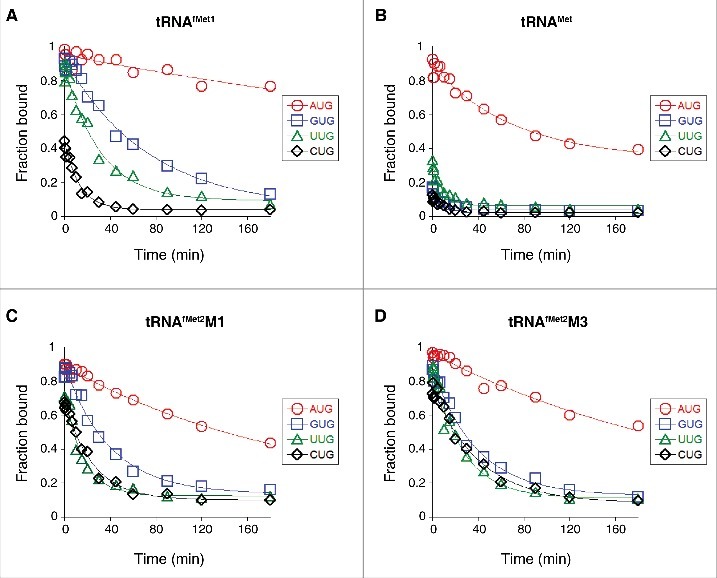
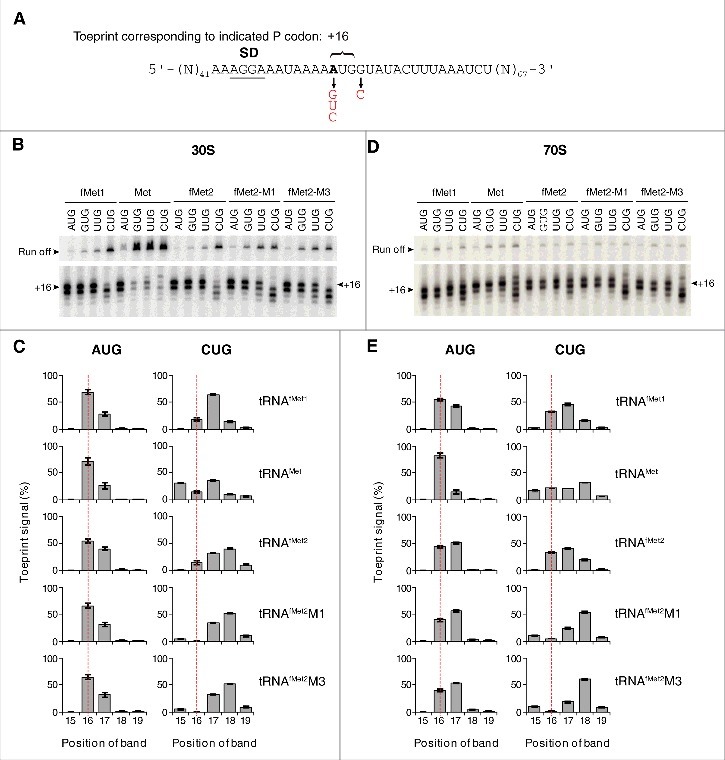
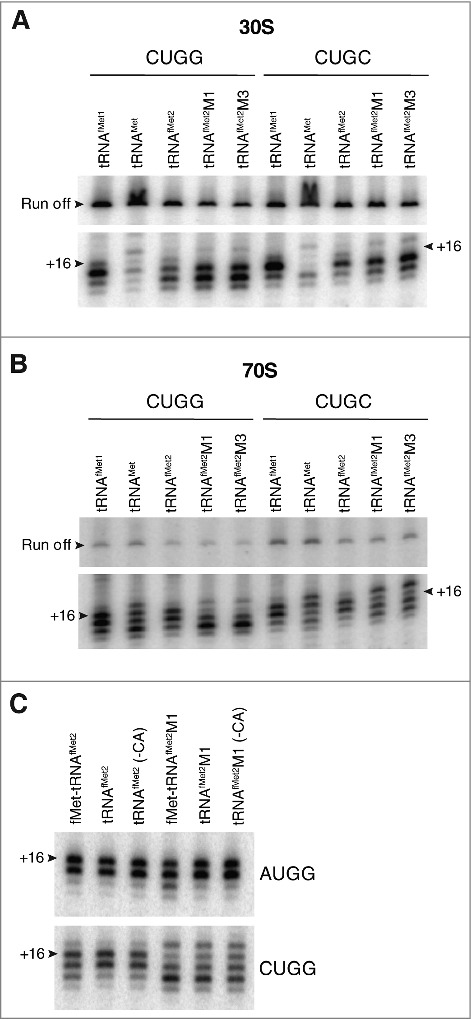
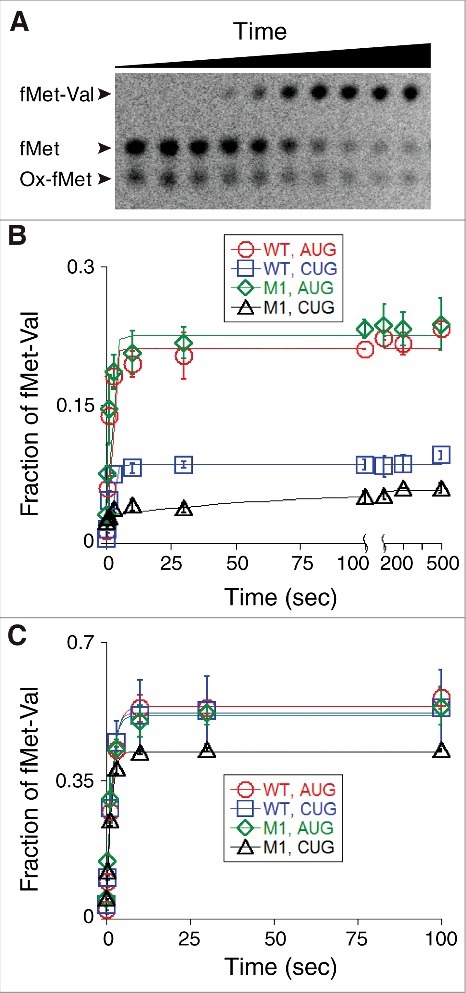
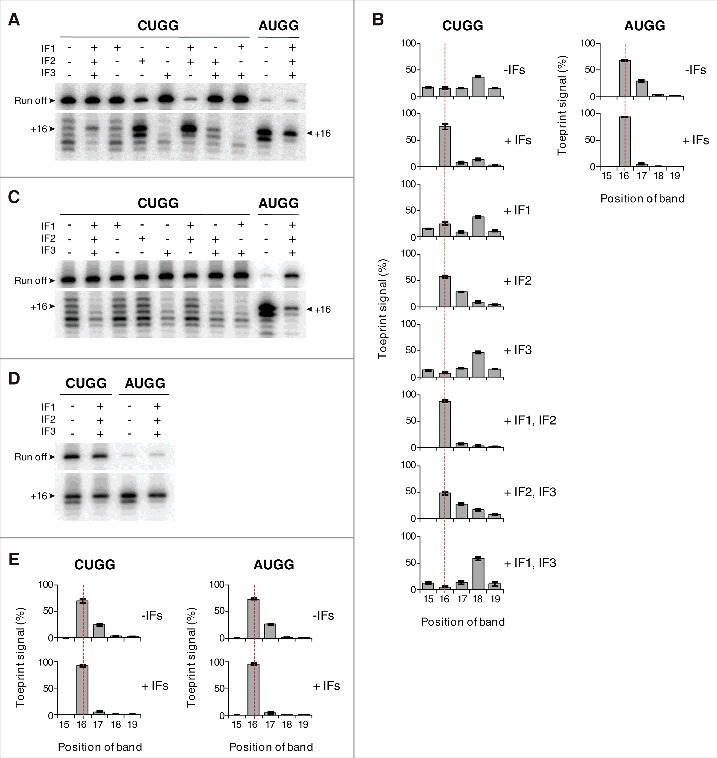
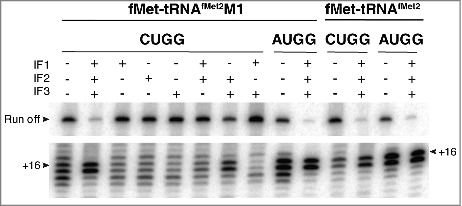
Translation begins at AUG, GUG, or UUG codons in bacteria. Start codon recognition occurs in the P site, which may help explain this first-position degeneracy. However, the molecular basis of start codon specificity remains unclear. In this study, we measured the codon dependence of 30S•mRNA•tRNAfMet and 30S•mRNA•tRNAMet complex formation. We found that complex stability varies over a large range with initiator tRNAfMet, following the same trend as reported previously for initiation rate in vivo (AUG > GUG, UUG > CUG, AUC, AUA > ACG). With elongator tRNAMet, the codon dependence of binding differs qualitatively, with virtually no discrimination between GUG and CUG. A unique feature of initiator tRNAfMet is a series of three G-C basepairs in the anticodon stem, which are known to be important for efficient initiation in vivo. A mutation targeting the central of these G-C basepairs causes the mRNA binding specificity pattern to change in a way reminiscent of elongator tRNAMet. Unexpectedly, for certain complexes containing fMet-tRNAfMet, we observed mispositioning of mRNA, such that codon 2 is no longer programmed in the A site. This mRNA mispositioning is exacerbated by the anticodon stem mutation and suppressed by IF2. These findings suggest that both IF2 and the unique anticodon stem of fMet-tRNAfMet help constrain mRNA positioning to set the correct reading frame during initiation.
Keywords: IF1; IF3; P site; initiation; ribosome; start codon selection.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous