

Resistance to CTLA-4 checkpoint inhibition reversed through selective elimination of granulocytic myeloid cells
- PMID: 28915554
- PMCID: PMC5593525
- DOI: 10.18632/oncotarget.18437
Resistance to CTLA-4 checkpoint inhibition reversed through selective elimination of granulocytic myeloid cells
Abstract
Purpose: Local immunosuppression remains a critical problem that limits clinically meaningful response to checkpoint inhibition in patients with head and neck cancer. Here, we assessed the impact of MDSC elimination on responses to CTLA-4 checkpoint inhibition.
Experimental design: Murine syngeneic carcinoma immune infiltrates were characterized by flow cytometry. Granulocytic MDSCs (gMDSCs) were depleted and T-lymphocyte antigen-specific responses were measured. Tumor-bearing mice were treated with MDSC depletion and CTLA-4 checkpoint blockade. Immune signatures within the human HNSCC datasets from The Cancer Genome Atlas (TCGA) were analyzed and differentially expressed genes from sorted human peripheral MDSCs were examined.
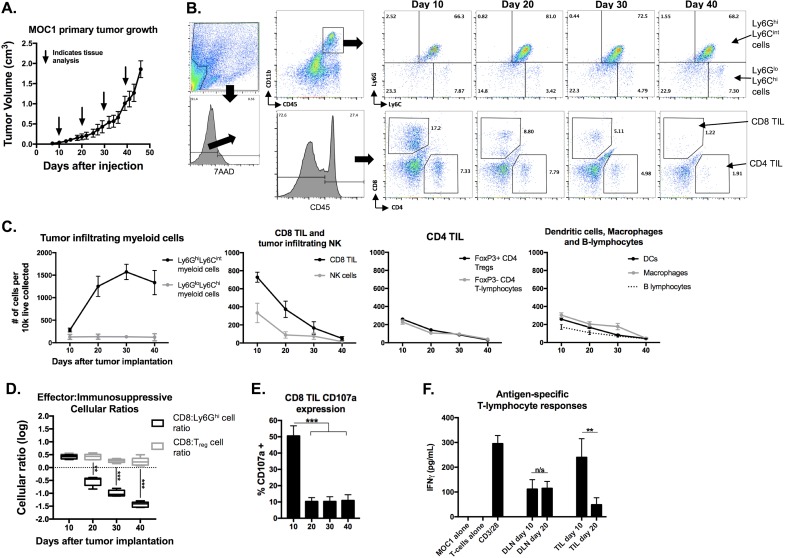
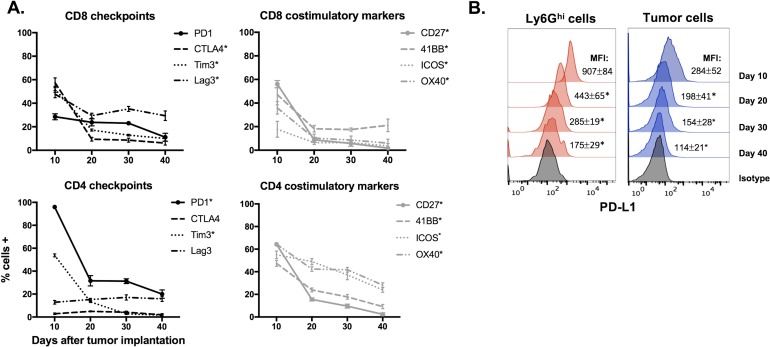
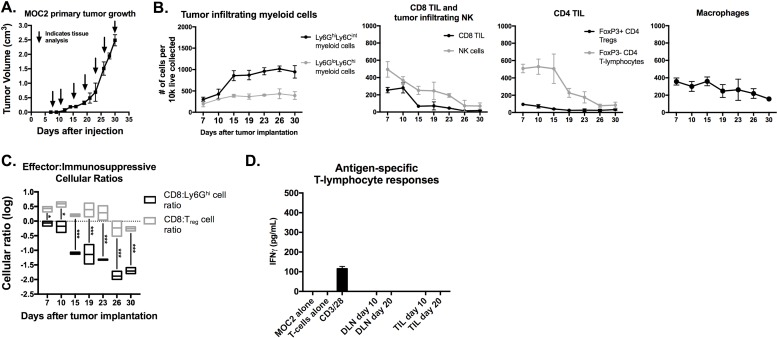
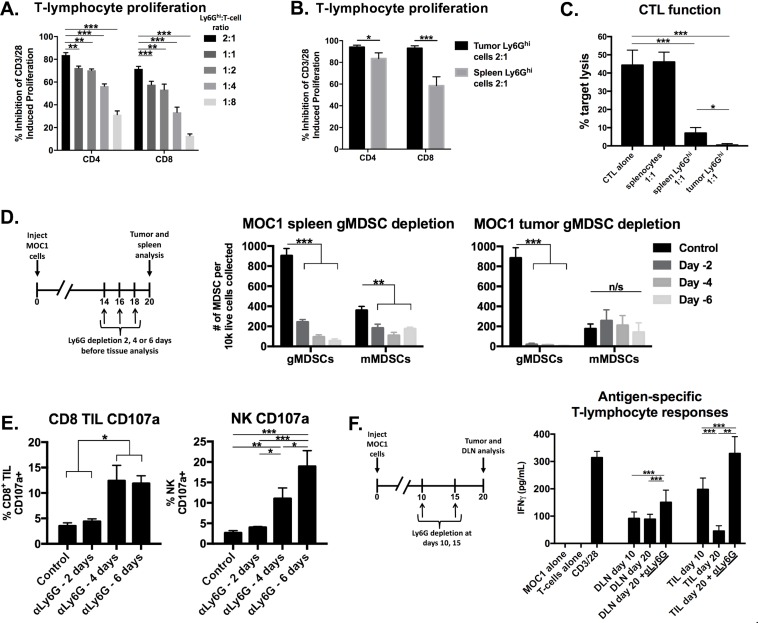
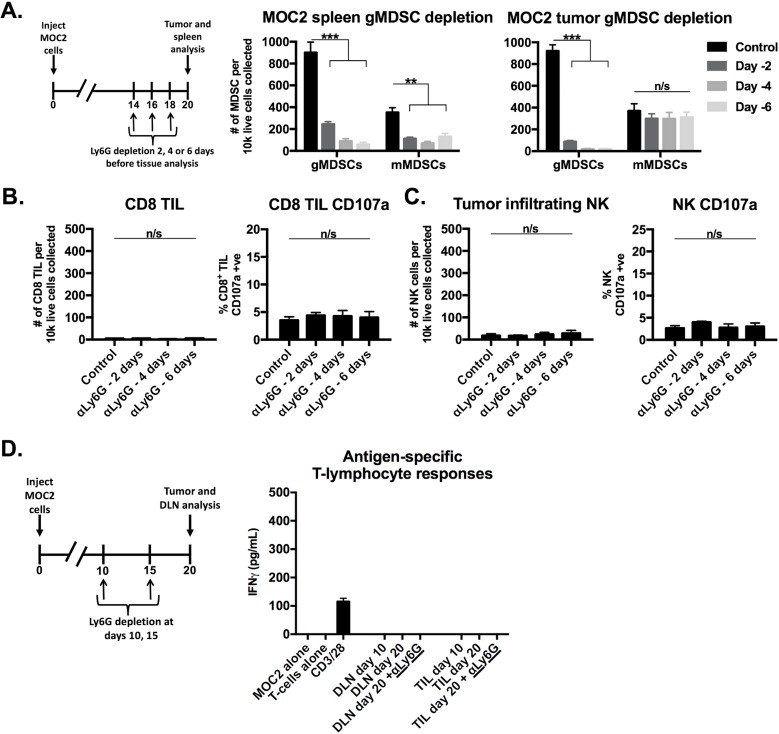
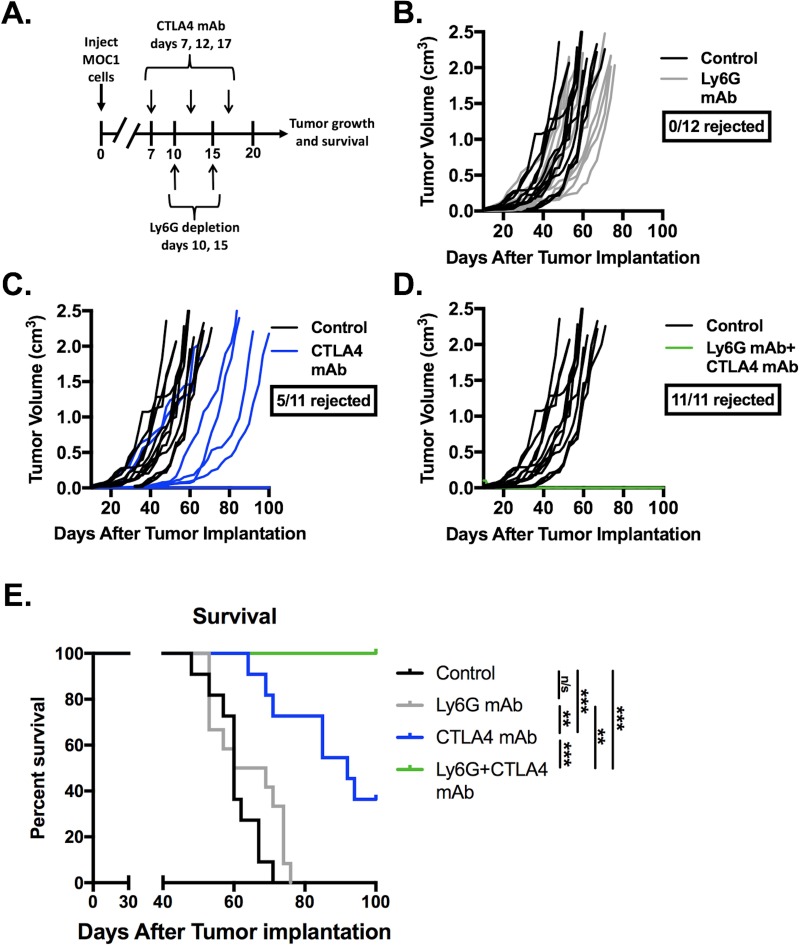
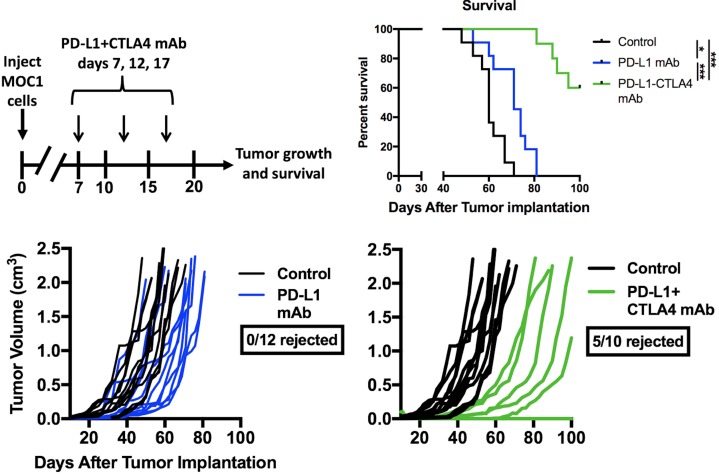
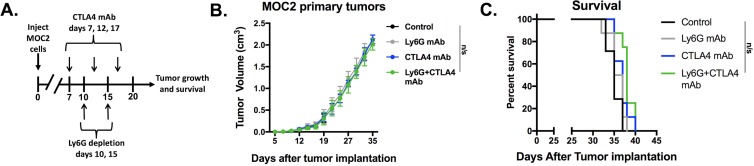
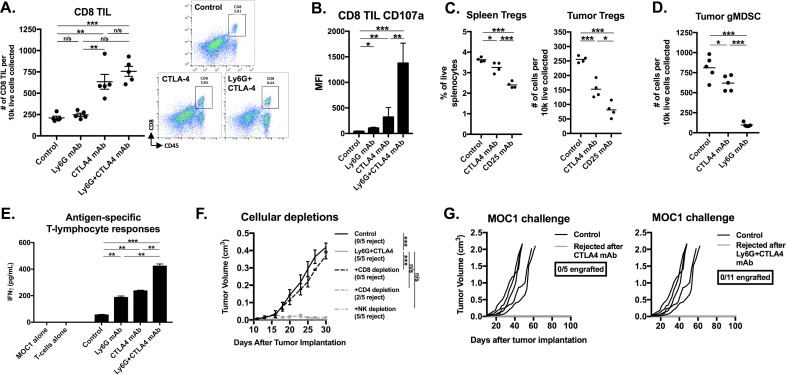
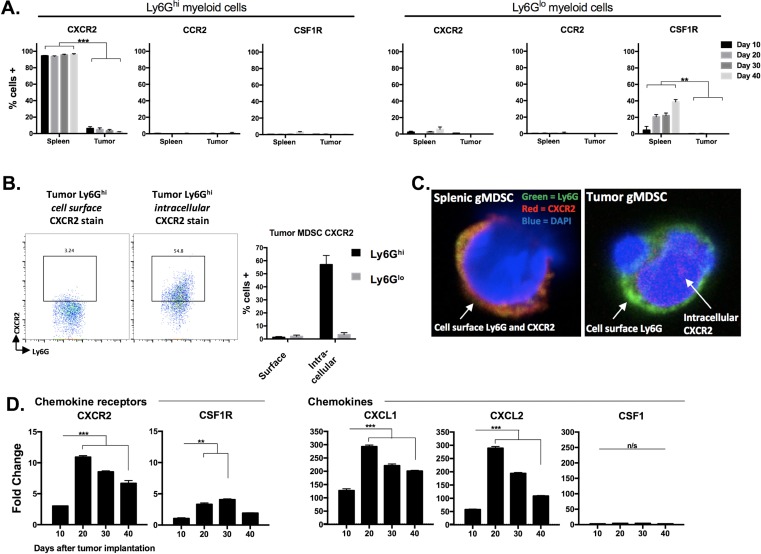
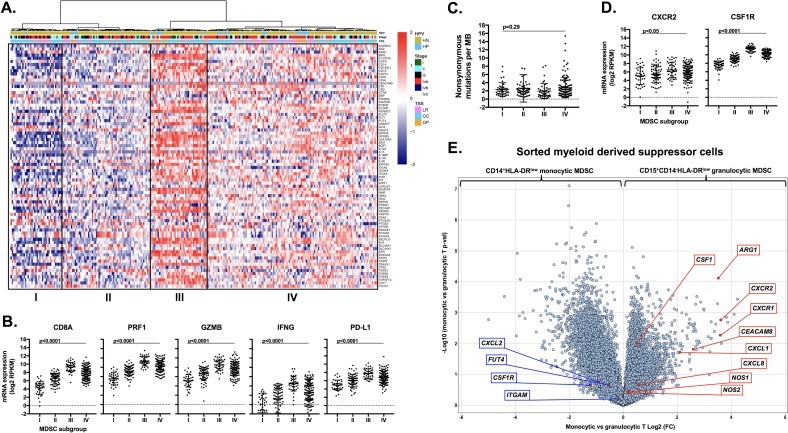
Results: gMDSCs accumulated with tumor progression and correlated with depletion of effector immune cells. Selective depletion of gMDSC restored tumor and draining lymph node antigen-specific T-lymphocyte responses lost with tumor progression. A subset of T-cell inflamed tumors responded to CTLA-4 mAb alone, but the addition of gMDSC depletion induced CD8 T-lymphocyte-dependent rejection of established tumors in all treated mice that resulted in immunologic memory. MDSCs differentially expressed chemokine receptors. Analysis of the head and neck cancer TCGA cohort revealed high CTLA-4 and MDSC-related chemokine and an MDSC-rich gene expression profile with a T-cell inflamed phenotype in > 60% of patients. CXCR2 and CSF1R expression was validated on sorted peripheral blood MDSCs from HNSCC patients.
Conclusions: MDSCs are a major contributor to local immunosuppression that limits responses to checkpoint inhibition in head and neck cancer. Limitation of MDSC recruitment or function represents a rational strategy to enhance responses to CTLA-4-based checkpoint inhibition in these patients.
Keywords: CTLA-4; MDSCs; T-cell inflamed; TCGA; rejection.
Conflict of interest statement
CONFLICTS OF INTEREST None.
Figures

References
-
- Schumacher TN, Schreiber RD. Neoantigens in cancer immunotherapy. Science. 2015;348:69–74. - PubMed
-
- Keck MK, Zuo Z, Khattri A, Stricker TP, Brown CD, Imanguli M, Rieke D, Endhardt K, Fang P, Brägelmann J, DeBoer R, El-Dinali M, Aktolga S, et al. Integrative analysis of head and neck cancer identifies two biologically distinct HPV and three non-HPV subtypes. Clin Cancer Res. 2015;21:870–81. - PubMed
-
- Seiwert TY, Burtness B, Mehra R, Weiss J, Berger R, Eder JP, Heath K, McClanahan T, Lunceford J, Gause C, Cheng JD, Chow LQ. Safety and clinical activity of pembrolizumab for treatment of recurrent or metastatic squamous cell carcinoma of the head and neck (KEYNOTE-012): an open-label, multicentre, phase 1b trial. Lancet Oncol. 2016;17:956–65. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
