

Down-regulation of NOX2 activity in phagocytes mediated by ATM-kinase dependent phosphorylation
- PMID: 28916473
- PMCID: PMC5699957
- DOI: 10.1016/j.freeradbiomed.2017.09.007
Down-regulation of NOX2 activity in phagocytes mediated by ATM-kinase dependent phosphorylation
Abstract
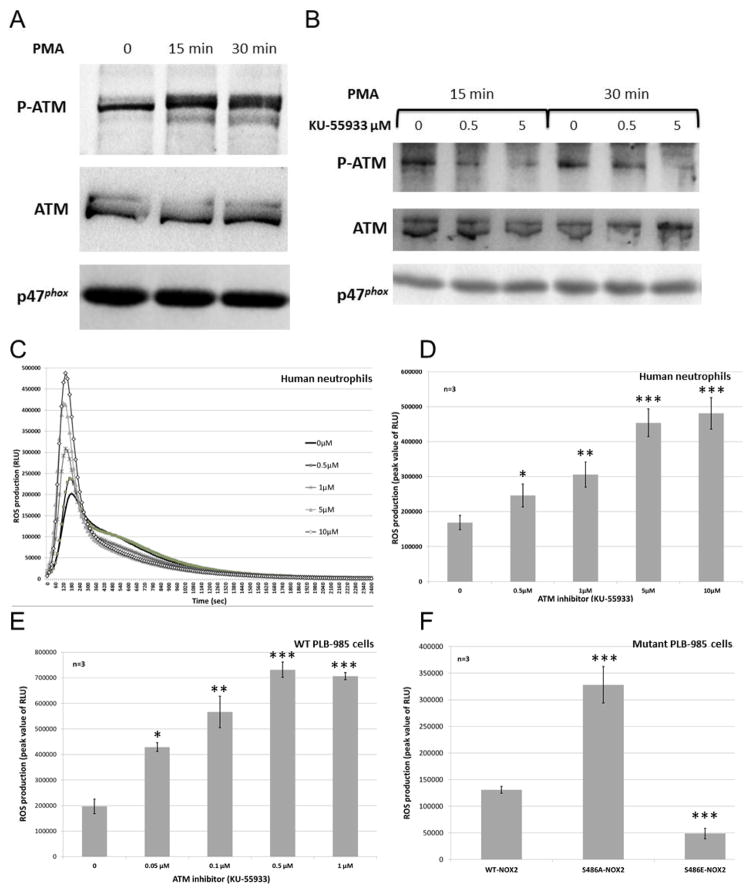
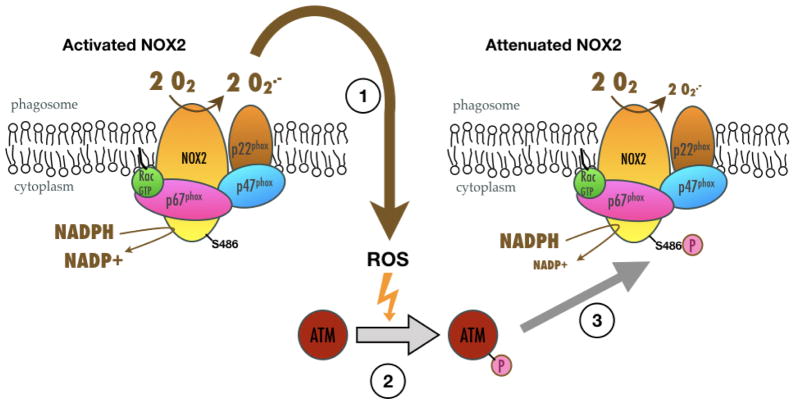
NADPH oxidases (NOX) have many biological roles, but their regulation to control production of potentially toxic ROS molecules remains unclear. A previously identified insertion sequence of 21 residues (called NIS) influences NOX activity, and its predicted flexibility makes it a good candidate for providing a dynamic switch controlling the NOX active site. We constructed NOX2 chimeras in which NIS had been deleted or exchanged with those from other NOXs (NIS1, 3 and 4). All contained functional heme and were expressed normally at the plasma membrane of differentiated PLB-985 cells. However, NOX2-ΔNIS and NOX2-NIS1 had neither NADPH-oxidase nor reductase activity and exhibited abnormal translocation of p47phox and p67phox to the phagosomal membrane. This suggested a functional role of NIS. Interestingly after activation, NOX2-NIS3 cells exhibited superoxide overproduction compared with wild-type cells. Paradoxically, the Vmax of purified unstimulated NOX2-NIS3 was only one-third of that of WT-NOX2. We therefore hypothesized that post-translational events regulate NOX2 activity and differ between NOX2-NIS3 and WT-NOX2. We demonstrated that Ser486, a phosphorylation target of ataxia telangiectasia mutated kinase (ATM kinase) located in the NIS of NOX2 (NOX2-NIS), was phosphorylated in purified cytochrome b558 after stimulation with phorbol 12-myristate-13-acetate (PMA). Moreover, ATM kinase inhibition and a NOX2 Ser486Ala mutation enhanced NOX activity whereas a Ser486Glu mutation inhibited it. Thus, the absence of Ser486 in NIS3 could explain the superoxide overproduction in the NOX2-NIS3 mutant. These results suggest that PMA-stimulated NOX2-NIS phosphorylation by ATM kinase causes a dynamic switch that deactivates NOX2 activity. We hypothesize that this downregulation is defective in NOX2-NIS3 mutant because of the absence of Ser486.
Keywords: Ataxia telangiectasia mutated (ATM); NADPH oxidase; NOX; NOX-specific Insertion Sequence (NIS); Neutrophil; Phosphorylation.
Copyright © 2017 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article.
Figures






Similar articles
-
Characterization of superoxide overproduction by the D-Loop(Nox4)-Nox2 cytochrome b(558) in phagocytes-Differential sensitivity to calcium and phosphorylation events.Biochim Biophys Acta. 2011 Jan;1808(1):78-90. doi: 10.1016/j.bbamem.2010.08.002. Epub 2010 Aug 11. Biochim Biophys Acta. 2011. PMID: 20708598 Free PMC article.
-
Regulation of the phagocyte NADPH oxidase activity: phosphorylation of gp91phox/NOX2 by protein kinase C enhances its diaphorase activity and binding to Rac2, p67phox, and p47phox.FASEB J. 2009 Apr;23(4):1011-22. doi: 10.1096/fj.08-114553. Epub 2008 Nov 21. FASEB J. 2009. PMID: 19028840 Free PMC article.
-
The protein kinase A negatively regulates reactive oxygen species production by phosphorylating gp91phox/NOX2 in human neutrophils.Free Radic Biol Med. 2020 Nov 20;160:19-27. doi: 10.1016/j.freeradbiomed.2020.07.021. Epub 2020 Aug 3. Free Radic Biol Med. 2020. PMID: 32758662
-
Strategies for identifying synthetic peptides to act as inhibitors of NADPH oxidases, or "all that you did and did not want to know about Nox inhibitory peptides".Cell Mol Life Sci. 2012 Jul;69(14):2283-305. doi: 10.1007/s00018-012-1007-4. Epub 2012 May 6. Cell Mol Life Sci. 2012. PMID: 22562603 Free PMC article. Review.
-
Role of the small GTPase Rac in p22phox-dependent NADPH oxidases.Biochimie. 2007 Sep;89(9):1133-44. doi: 10.1016/j.biochi.2007.05.003. Epub 2007 May 17. Biochimie. 2007. PMID: 17583407 Review.
Cited by
-
NADPH Oxidases (NOX): An Overview from Discovery, Molecular Mechanisms to Physiology and Pathology.Antioxidants (Basel). 2021 Jun 1;10(6):890. doi: 10.3390/antiox10060890. Antioxidants (Basel). 2021. PMID: 34205998 Free PMC article. Review.
-
CRK2 and C-terminal Phosphorylation of NADPH Oxidase RBOHD Regulate Reactive Oxygen Species Production in Arabidopsis.Plant Cell. 2020 Apr;32(4):1063-1080. doi: 10.1105/tpc.19.00525. Epub 2020 Feb 7. Plant Cell. 2020. PMID: 32034035 Free PMC article.
-
Inhibition of NADPH oxidase alleviates germ cell apoptosis and ER stress during testicular ischemia reperfusion injury.Saudi J Biol Sci. 2020 Aug;27(8):2174-2184. doi: 10.1016/j.sjbs.2020.04.024. Epub 2020 Apr 21. Saudi J Biol Sci. 2020. PMID: 32714044 Free PMC article.
-
The Emerging Roles of Nicotinamide Adenine Dinucleotide Phosphate Oxidase 2 in Skeletal Muscle Redox Signaling and Metabolism.Antioxid Redox Signal. 2019 Dec 20;31(18):1371-1410. doi: 10.1089/ars.2018.7678. Epub 2019 Nov 1. Antioxid Redox Signal. 2019. PMID: 31588777 Free PMC article. Review.
-
Clinical, functional and genetic characterization of 16 patients suffering from chronic granulomatous disease variants - identification of 11 novel mutations in CYBB.Clin Exp Immunol. 2021 Feb;203(2):247-266. doi: 10.1111/cei.13520. Epub 2020 Oct 12. Clin Exp Immunol. 2021. PMID: 32954498 Free PMC article.
References
-
- Bedard K, Krause KH. The NOX family of ROS-generating NADPH oxidases: physiology and pathophysiology. Physiol Rev. 2007;87:245–313. - PubMed
-
- Paravicini TM, Touyz RM. NADPH oxidases, reactive oxygen species, and hypertension: clinical implications and therapeutic possibilities. Diabetes Care. 2008;31(Suppl 2):S170–180. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
