

Hormetic potential of methylglyoxal, a side-product of glycolysis, in switching tumours from growth to death
- PMID: 28916747
- PMCID: PMC5600983
- DOI: 10.1038/s41598-017-12119-7
Hormetic potential of methylglyoxal, a side-product of glycolysis, in switching tumours from growth to death
Abstract
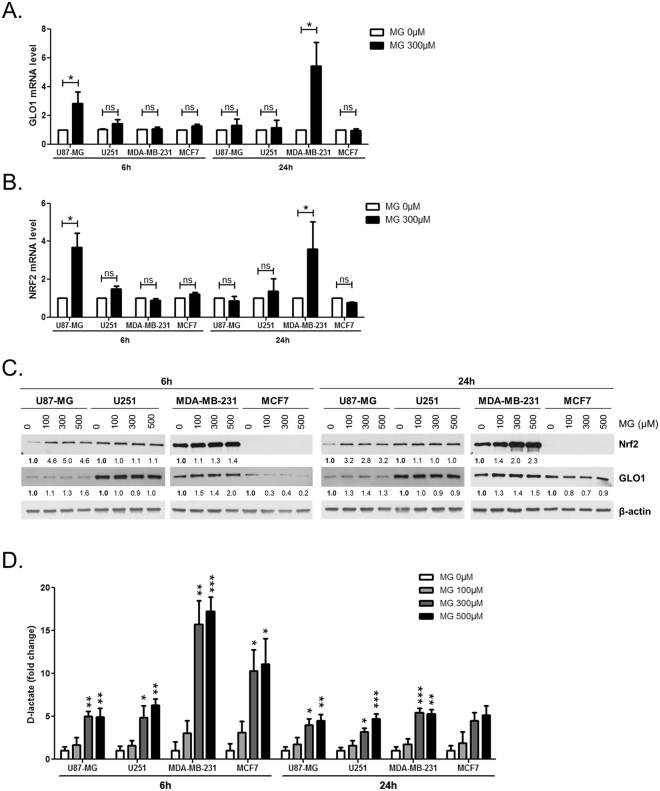
Metabolic reprogramming toward aerobic glycolysis unavoidably favours methylglyoxal (MG) and advanced glycation end products (AGEs) formation in cancer cells. MG was initially considered a highly cytotoxic molecule with potential anti-cancer value. However, we have recently demonstrated that MG enhanced tumour growth and metastasis. In an attempt to understand this dual role, we explored MG-mediated dicarbonyl stress status in four breast and glioblastoma cancer cell lines in relation with their glycolytic phenotype and MG detoxifying capacity. In glycolytic cancer cells cultured in high glucose, we observed a significant increase of the conversion of MG to D-lactate through the glyoxalase system. Moreover, upon exogenous MG challenge, glycolytic cells showed elevated amounts of intracellular MG and induced de novo GLO1 detoxifying enzyme and Nrf2 expression. Thus, supporting the adaptive nature of glycolytic cancer cells to MG dicarbonyl stress when compared to non-glycolytic ones. Finally and consistent with the pro-tumoural role of MG, we showed that low doses of MG induced AGEs formation and tumour growth in vivo, both of which can be reversed using a MG scavenger. Our study represents the first demonstration of a hormetic effect of MG defined by a low-dose stimulation and a high-dose inhibition of tumour growth.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures






References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
