

Ampk phosphorylation of Ulk1 is required for targeting of mitochondria to lysosomes in exercise-induced mitophagy
- PMID: 28916822
- PMCID: PMC5601463
- DOI: 10.1038/s41467-017-00520-9
Ampk phosphorylation of Ulk1 is required for targeting of mitochondria to lysosomes in exercise-induced mitophagy
Abstract
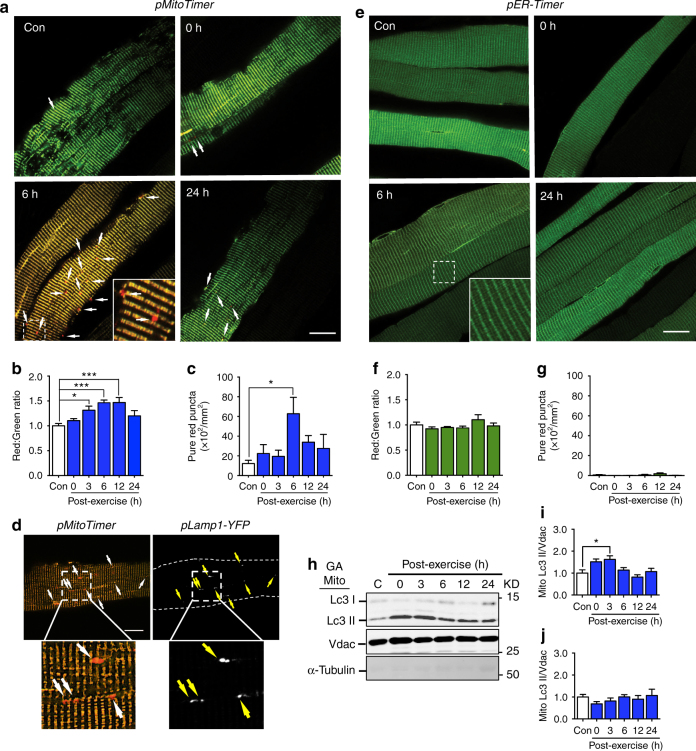
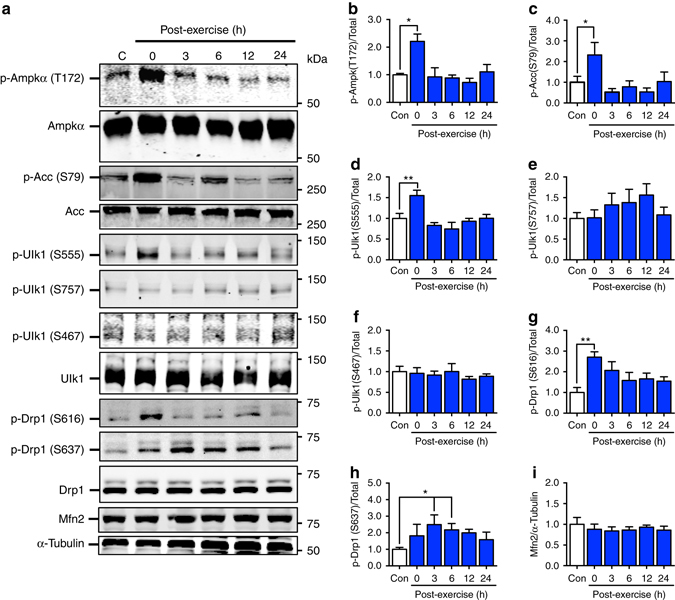
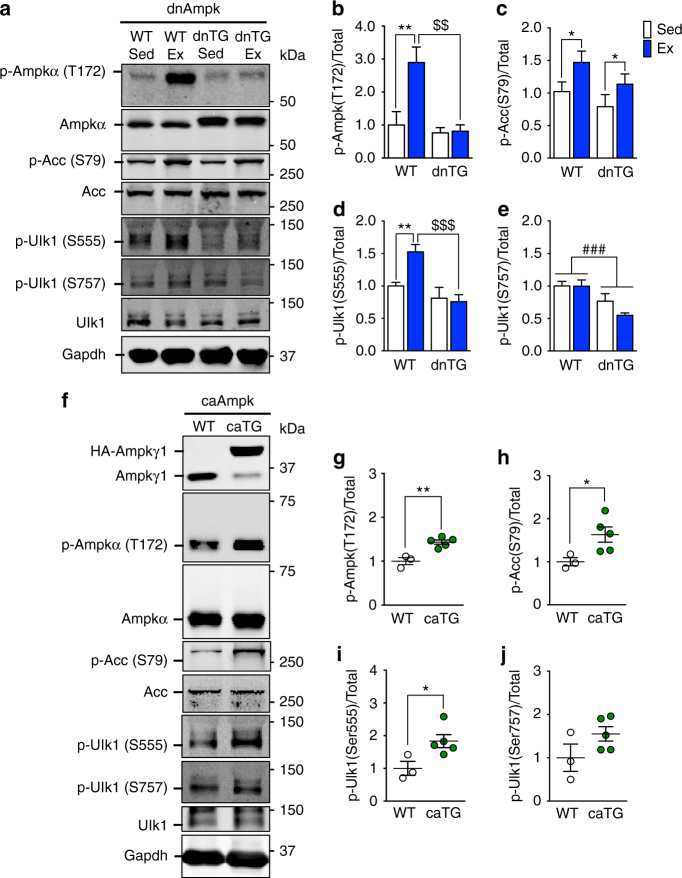
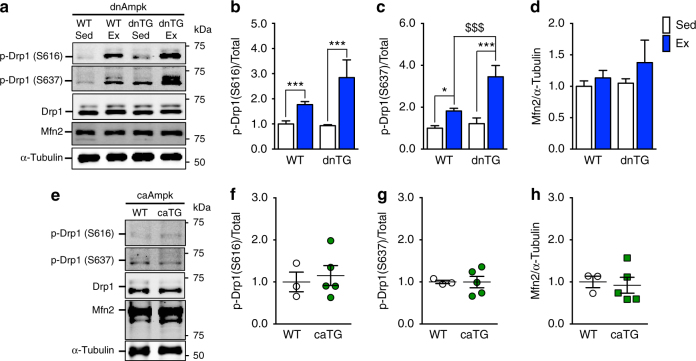
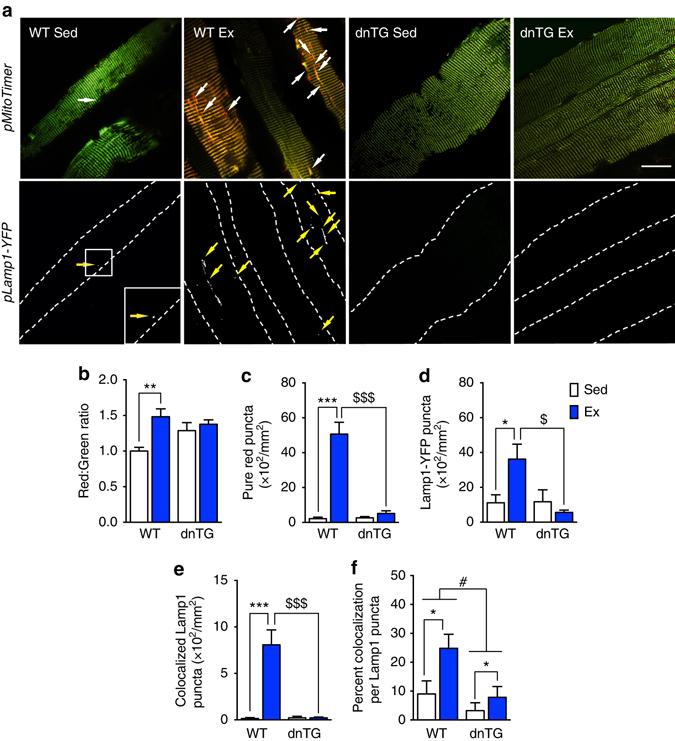
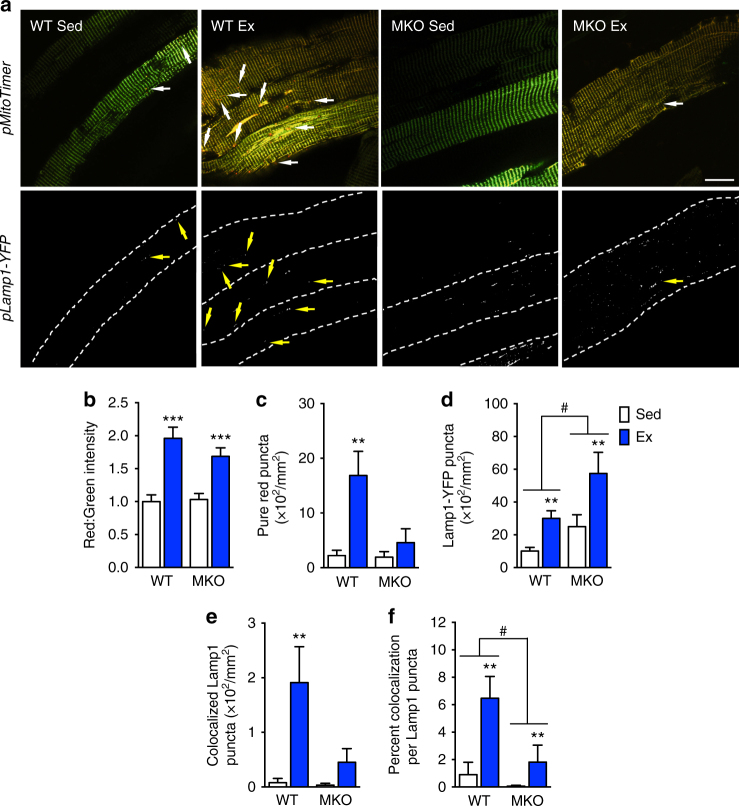
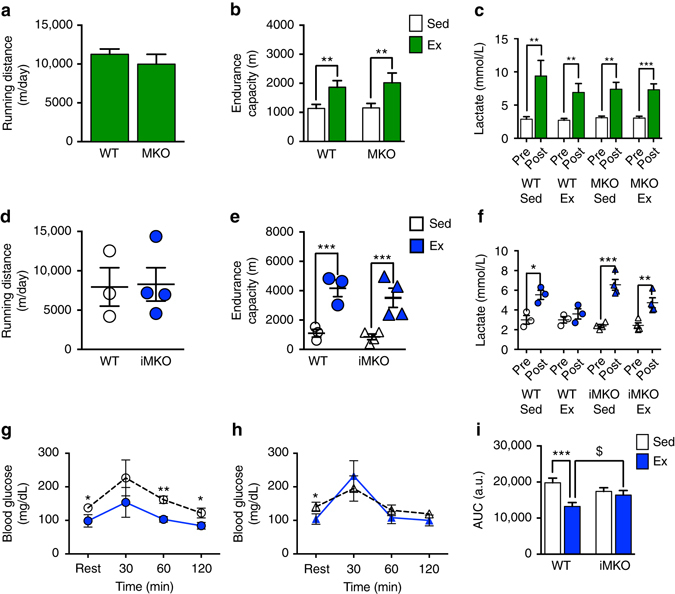
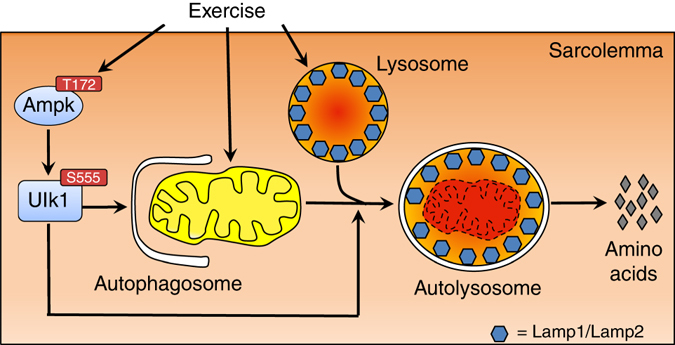
Mitochondrial health is critical for skeletal muscle function and is improved by exercise training through both mitochondrial biogenesis and removal of damaged/dysfunctional mitochondria via mitophagy. The mechanisms underlying exercise-induced mitophagy have not been fully elucidated. Here, we show that acute treadmill running in mice causes mitochondrial oxidative stress at 3-12 h and mitophagy at 6 h post-exercise in skeletal muscle. These changes were monitored using a novel fluorescent reporter gene, pMitoTimer, that allows assessment of mitochondrial oxidative stress and mitophagy in vivo, and were preceded by increased phosphorylation of AMP activated protein kinase (Ampk) at tyrosine 172 and of unc-51 like autophagy activating kinase 1 (Ulk1) at serine 555. Using mice expressing dominant negative and constitutively active Ampk in skeletal muscle, we demonstrate that Ulk1 activation is dependent on Ampk. Furthermore, exercise-induced metabolic adaptation requires Ulk1. These findings provide direct evidence of exercise-induced mitophagy and demonstrate the importance of Ampk-Ulk1 signaling in skeletal muscle.Exercise is associated with biogenesis and removal of dysfunctional mitochondria. Here the authors use a mitochondrial reporter gene to demonstrate the occurrence of mitophagy following exercise in mice, and show this is dependent on AMPK and ULK1 signaling.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
Thyroid hormone induction of mitochondrial activity is coupled to mitophagy via ROS-AMPK-ULK1 signaling.Autophagy. 2015;11(8):1341-57. doi: 10.1080/15548627.2015.1061849. Autophagy. 2015. PMID: 26103054 Free PMC article.
-
Phosphorylation of ULK1 by AMPK regulates translocation of ULK1 to mitochondria and mitophagy.FEBS Lett. 2015 Jul 8;589(15):1847-54. doi: 10.1016/j.febslet.2015.05.020. Epub 2015 May 14. FEBS Lett. 2015. PMID: 25980607
-
Epimedii Folium and Ligustri Lucidi Fructus synergistically delay renal aging through AMPK/ULK1/Bcl2L13-mediated mitophagy.J Ethnopharmacol. 2025 Apr 25;346:119668. doi: 10.1016/j.jep.2025.119668. Epub 2025 Mar 21. J Ethnopharmacol. 2025. PMID: 40122318
-
Mitophagy: Molecular Mechanisms, New Concepts on Parkin Activation and the Emerging Role of AMPK/ULK1 Axis.Cells. 2021 Dec 23;11(1):30. doi: 10.3390/cells11010030. Cells. 2021. PMID: 35011593 Free PMC article. Review.
-
Exercise-mediated modulation of autophagy in skeletal muscle.Scand J Med Sci Sports. 2018 Mar;28(3):772-781. doi: 10.1111/sms.12945. Epub 2017 Aug 4. Scand J Med Sci Sports. 2018. PMID: 28685860 Review.
Cited by
-
Dexmedetomidine Protects Human Cardiomyocytes Against Ischemia-Reperfusion Injury Through α2-Adrenergic Receptor/AMPK-Dependent Autophagy.Front Pharmacol. 2021 May 21;12:615424. doi: 10.3389/fphar.2021.615424. eCollection 2021. Front Pharmacol. 2021. PMID: 34093174 Free PMC article.
-
Mitochondrial quality control in health and cardiovascular diseases.Front Cell Dev Biol. 2023 Nov 6;11:1290046. doi: 10.3389/fcell.2023.1290046. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 38020895 Free PMC article. Review.
-
TBC1D23 mediates Golgi-specific LKB1 signaling.Nat Commun. 2024 Feb 27;15(1):1785. doi: 10.1038/s41467-024-46166-2. Nat Commun. 2024. PMID: 38413626 Free PMC article.
-
Exercise and mitochondrial mechanisms in patients with sarcopenia.Front Physiol. 2022 Dec 6;13:1040381. doi: 10.3389/fphys.2022.1040381. eCollection 2022. Front Physiol. 2022. PMID: 36561214 Free PMC article. Review.
-
Skeletal muscle mitochondrial remodeling in exercise and diseases.Cell Res. 2018 Oct;28(10):969-980. doi: 10.1038/s41422-018-0078-7. Epub 2018 Aug 14. Cell Res. 2018. PMID: 30108290 Free PMC article. Review.
References
-
- Hollozy JO. Biochemical adaptations in muscle. Effects of exercise on mitochondrial oxygen uptake and respiratory enzyme activity in skeletal muscle. J. Biol. Chem. 1967;242:2278–2282. - PubMed
-
- Tseng BS, Marsh DR, Hamilton MT, Booth FW. Strength and aerobic training attenuate muscle wasting and improve resistance to the development of disability with aging. Cell Signal. 2015;50:113–119. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
