

Topological Structure Determination of RNA Using Small-Angle X-Ray Scattering
- PMID: 28918093
- PMCID: PMC5693684
- DOI: 10.1016/j.jmb.2017.09.006
Topological Structure Determination of RNA Using Small-Angle X-Ray Scattering
Abstract
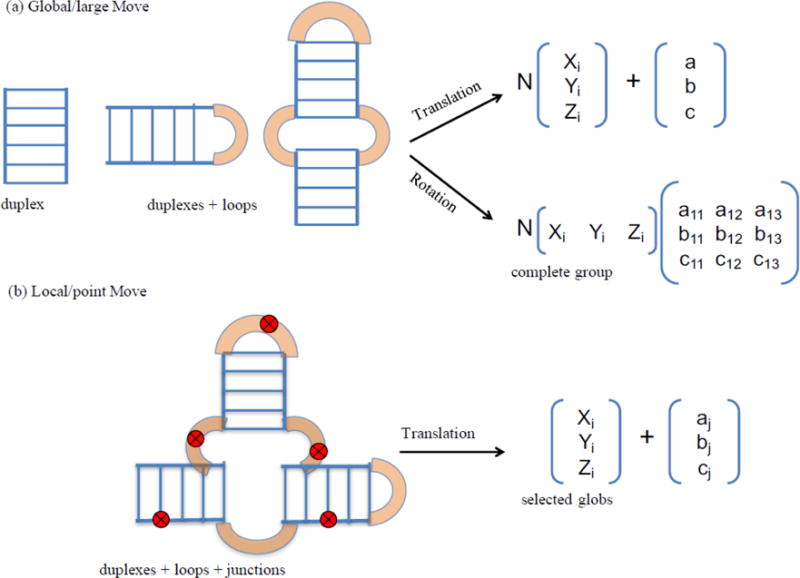
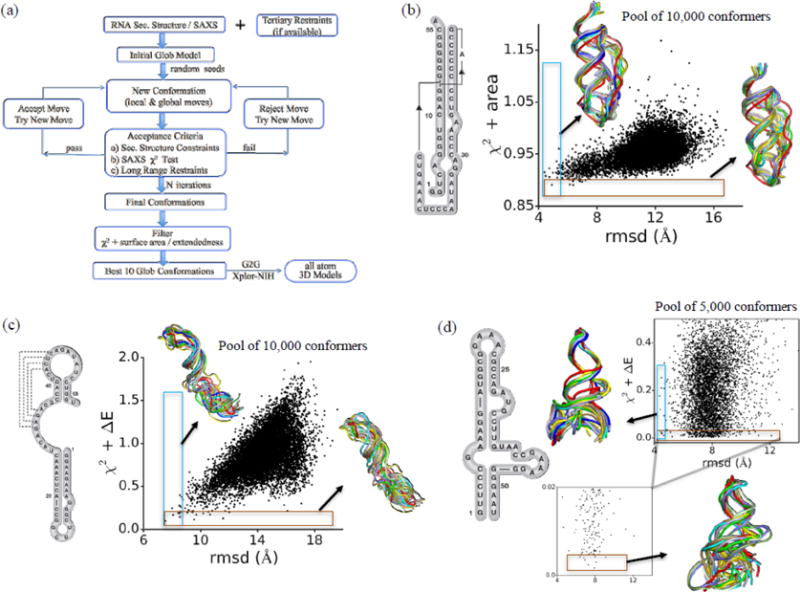
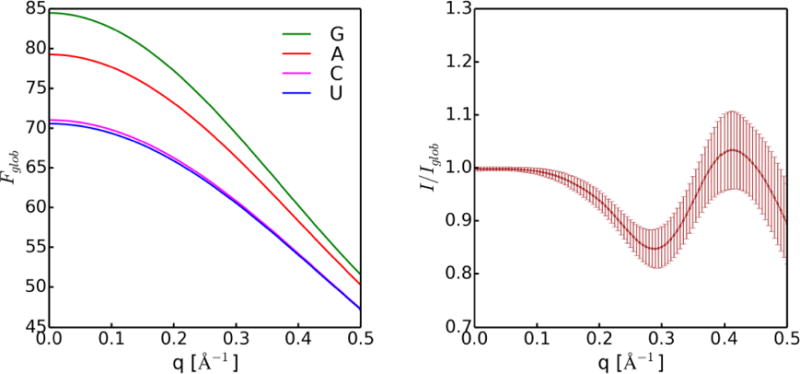
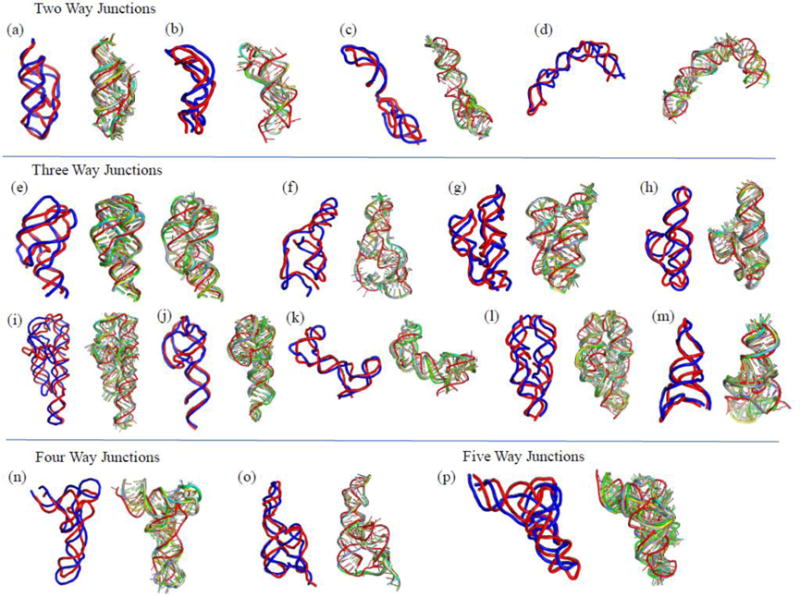
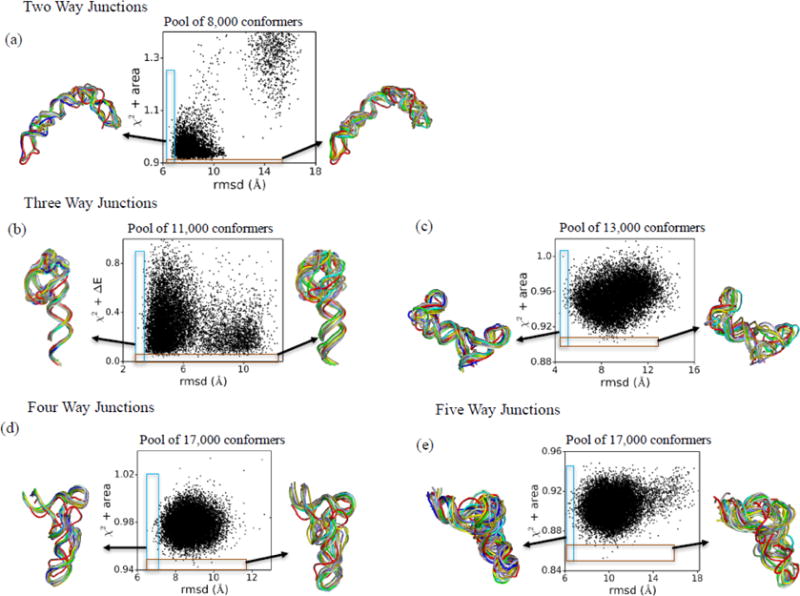
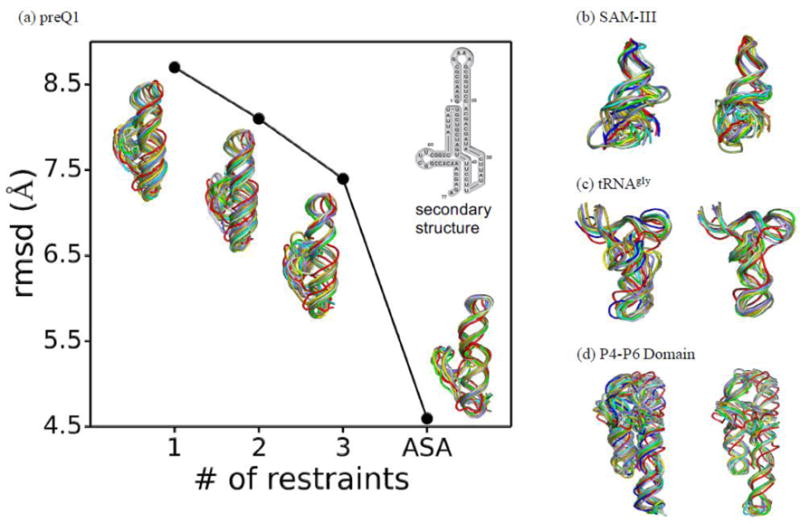
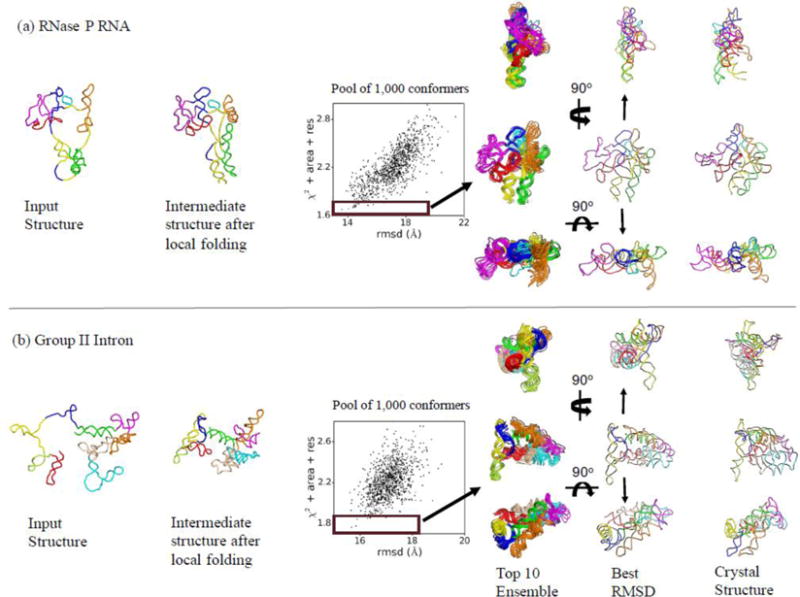
Knowledge of RNA three-dimensional topological structures provides important insight into the relationship between RNA structural components and function. It is often likely that near-complete sets of biochemical and biophysical data containing structural restraints are not available, but one still wants to obtain knowledge about approximate topological folding of RNA. In this regard, general methods for determining such topological structures with minimum readily available restraints are lacking. Naked RNAs are difficult to crystallize and NMR spectroscopy is generally limited to small RNA fragments. By nature, sequence determines structure and all interactions that drive folding are self-contained within sequence. Nevertheless, there is little apparent correlation between primary sequences and three-dimensional folding unless supplemented with experimental or phylogenetic data. Thus, there is an acute need for a robust high-throughput method that can rapidly determine topological structures of RNAs guided by some experimental data. We present here a novel method (RS3D) that can assimilate the RNA secondary structure information, small-angle X-ray scattering data, and any readily available tertiary contact information to determine the topological fold of RNA. Conformations are firstly sampled at glob level where each glob represents a nucleotide. Best-ranked glob models can be further refined against solvent accessibility data, if available, and then converted to explicit all-atom coordinates for refinement against SAXS data using the Xplor-NIH program. RS3D is widely applicable to a variety of RNA folding architectures currently present in the structure database. Furthermore, we demonstrate applicability and feasibility of the program to derive low-resolution topological structures of relatively large multi-domain RNAs.
Keywords: RNA secondary and tertiary structure; RNA topological structure; hierarchical moves; small-angle X-ray scattering.
Published by Elsevier Ltd.
Figures
Similar articles
-
Modeling RNA topological structures using small angle X-ray scattering.Methods. 2016 Jul 1;103:18-24. doi: 10.1016/j.ymeth.2016.04.015. Epub 2016 Jun 2. Methods. 2016. PMID: 27090001 Review.
-
Small-angle X-ray scattering: a bridge between RNA secondary structures and three-dimensional topological structures.Curr Opin Struct Biol. 2015 Feb;30:147-160. doi: 10.1016/j.sbi.2015.02.010. Epub 2015 Mar 10. Curr Opin Struct Biol. 2015. PMID: 25765781 Free PMC article. Review.
-
Deriving RNA topological structure from SAXS.Methods Enzymol. 2022;677:479-529. doi: 10.1016/bs.mie.2022.08.037. Epub 2022 Oct 26. Methods Enzymol. 2022. PMID: 36410961
-
Combined Small-Angle X-ray and Neutron Scattering Restraints in Molecular Dynamics Simulations.J Chem Theory Comput. 2019 Aug 13;15(8):4687-4698. doi: 10.1021/acs.jctc.9b00292. Epub 2019 Jul 30. J Chem Theory Comput. 2019. PMID: 31251056
-
RNA structure determination using SAXS data.J Phys Chem B. 2010 Aug 12;114(31):10039-48. doi: 10.1021/jp1057308. J Phys Chem B. 2010. PMID: 20684627 Free PMC article.
Cited by
-
Advances in RNA 3D Structure Modeling Using Experimental Data.Front Genet. 2020 Oct 26;11:574485. doi: 10.3389/fgene.2020.574485. eCollection 2020. Front Genet. 2020. PMID: 33193680 Free PMC article. Review.
-
Dynamic Structural Biology Experiments at XFEL or Synchrotron Sources.Methods Mol Biol. 2021;2305:203-228. doi: 10.1007/978-1-0716-1406-8_11. Methods Mol Biol. 2021. PMID: 33950392 Review.
-
Advances and opportunities in RNA structure experimental determination and computational modeling.Nat Methods. 2022 Oct;19(10):1193-1207. doi: 10.1038/s41592-022-01623-y. Epub 2022 Oct 6. Nat Methods. 2022. PMID: 36203019 Review.
-
Visualizing RNA conformational and architectural heterogeneity in solution.Nat Commun. 2023 Feb 9;14(1):714. doi: 10.1038/s41467-023-36184-x. Nat Commun. 2023. PMID: 36759615 Free PMC article.
-
Determining structures of RNA conformers using AFM and deep neural networks.Nature. 2025 Jan;637(8048):1234-1243. doi: 10.1038/s41586-024-07559-x. Epub 2024 Dec 18. Nature. 2025. PMID: 39695231 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
