

Adaptive Evolution Leads to Cross-Species Incompatibility in the piRNA Transposon Silencing Machinery
- PMID: 28919205
- PMCID: PMC5653967
- DOI: 10.1016/j.devcel.2017.08.012
Adaptive Evolution Leads to Cross-Species Incompatibility in the piRNA Transposon Silencing Machinery
Abstract
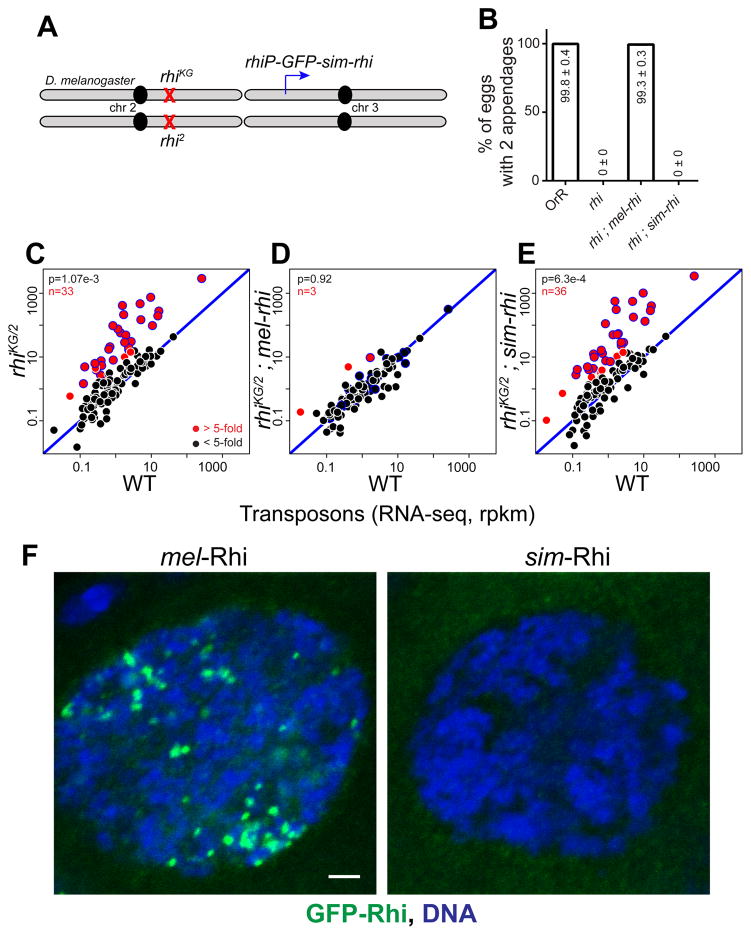
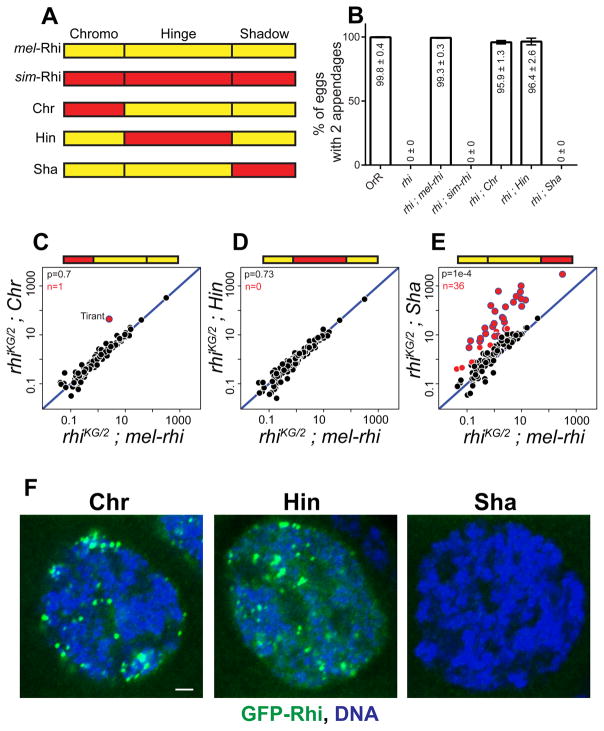
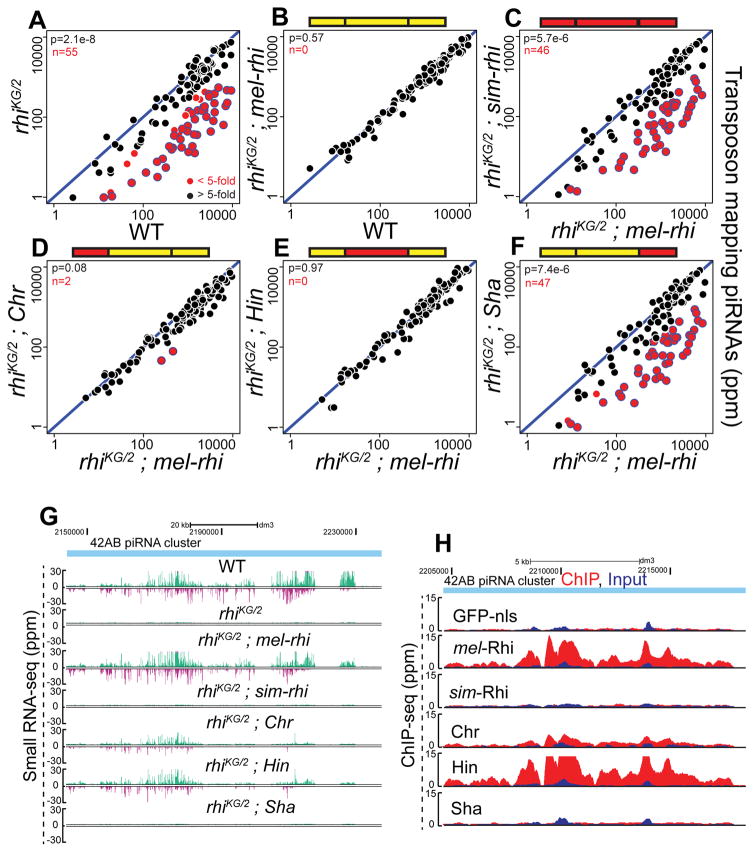
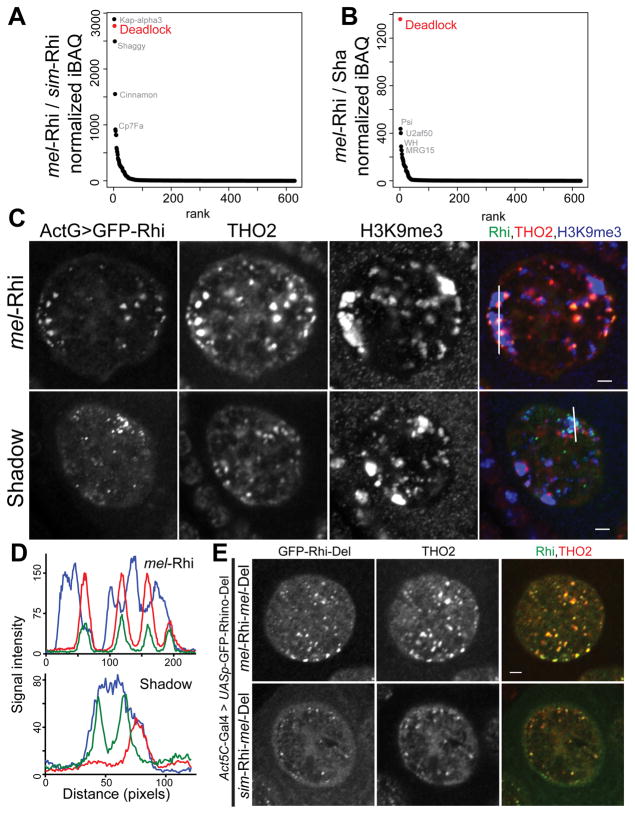
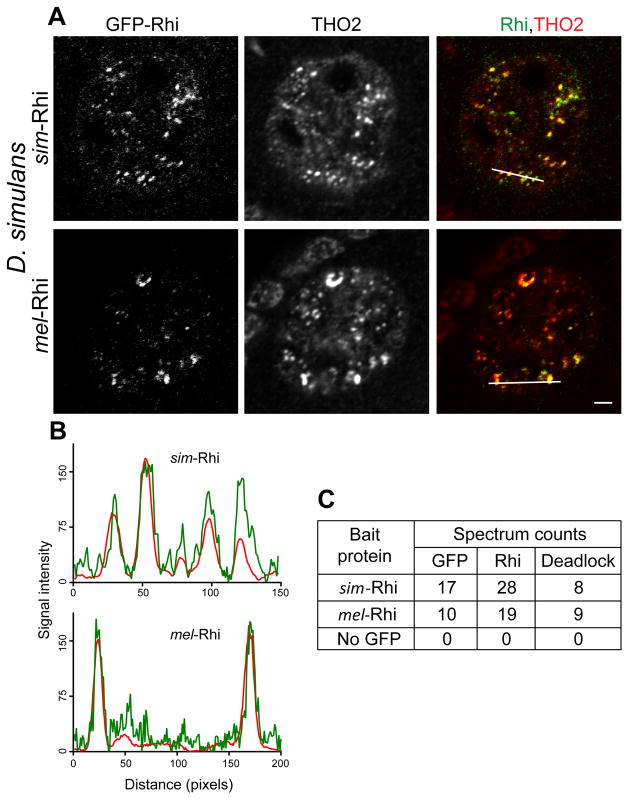
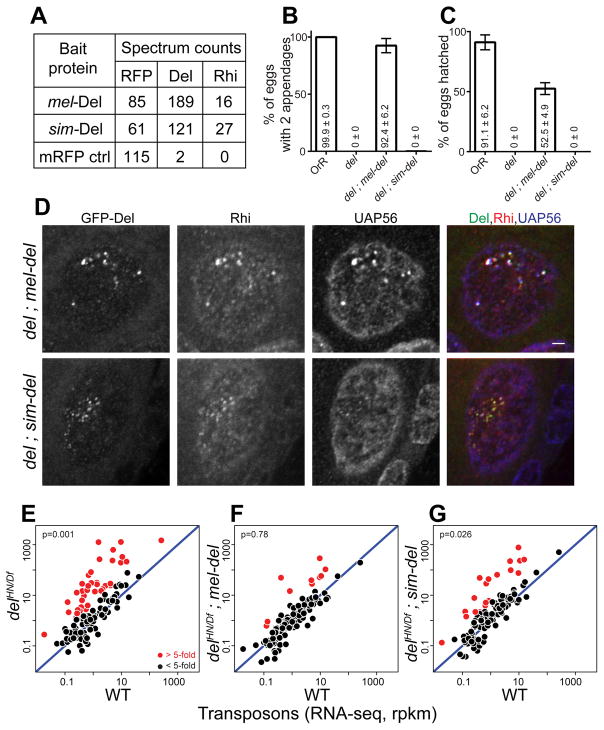
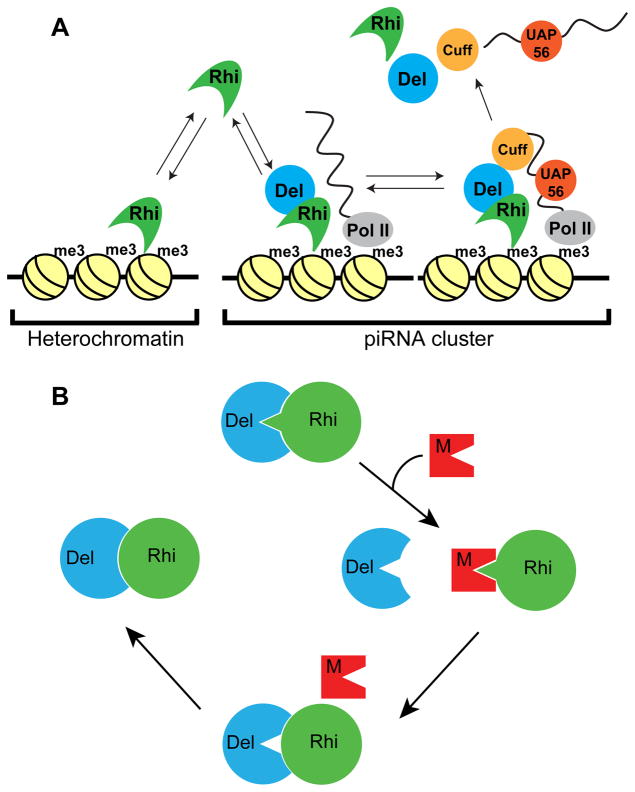
Reproductive isolation defines species divergence and is linked to adaptive evolution of hybrid incompatibility genes. Hybrids between Drosophila melanogaster and Drosophila simulans are sterile, and phenocopy mutations in the PIWI interacting RNA (piRNA) pathway, which silences transposons and shows pervasive adaptive evolution, and Drosophila rhino and deadlock encode rapidly evolving components of a complex that binds to piRNA clusters. We show that Rhino and Deadlock interact and co-localize in simulans and melanogaster, but simulans Rhino does not bind melanogaster Deadlock, due to substitutions in the rapidly evolving Shadow domain. Significantly, a chimera expressing the simulans Shadow domain in a melanogaster Rhino backbone fails to support piRNA production, disrupts binding to piRNA clusters, and leads to ectopic localization to bulk heterochromatin. Fusing melanogaster Deadlock to simulans Rhino, by contrast, restores localization to clusters. Deadlock binding thus directs Rhino to piRNA clusters, and Rhino-Deadlock co-evolution has produced cross-species incompatibilities, which may contribute to reproductive isolation.
Keywords: adaptive evolution; chromatin; piRNA; reproductive isolation; transposon silencing.
Copyright © 2017. Published by Elsevier Inc.
Figures
Comment in
-
Germline Genome Protection on an Evolutionary Treadmill.Dev Cell. 2017 Oct 9;43(1):1-3. doi: 10.1016/j.devcel.2017.09.021. Dev Cell. 2017. PMID: 29017026
References
-
- Bannister AJ, Zegerman P, Partridge JF, Miska EA, Thomas JO, Allshire RC, Kouzarides T. Selective recognition of methylated lysine 9 on histone H3 by the HP1 chromo domain. Nature. 2001;410:120–124. - PubMed
-
- Belancio VP, Hedges DJ, Deininger P. Mammalian non-LTR retrotransposons: for better or worse, in sickness and in health. Genome Res. 2008;18:343–358. - PubMed
-
- Belyayev A. Bursts of transposable elements as an evolutionary driving force. J Evol Biol. 2014;27:2573–2584. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
