

Heterotrimeric G Protein-Regulated Ca2+ Influx and PIN2 Asymmetric Distribution Are Involved in Arabidopsis thaliana Roots' Avoidance Response to Extracellular ATP
- PMID: 28919907
- PMCID: PMC5585194
- DOI: 10.3389/fpls.2017.01522
Heterotrimeric G Protein-Regulated Ca2+ Influx and PIN2 Asymmetric Distribution Are Involved in Arabidopsis thaliana Roots' Avoidance Response to Extracellular ATP
Abstract
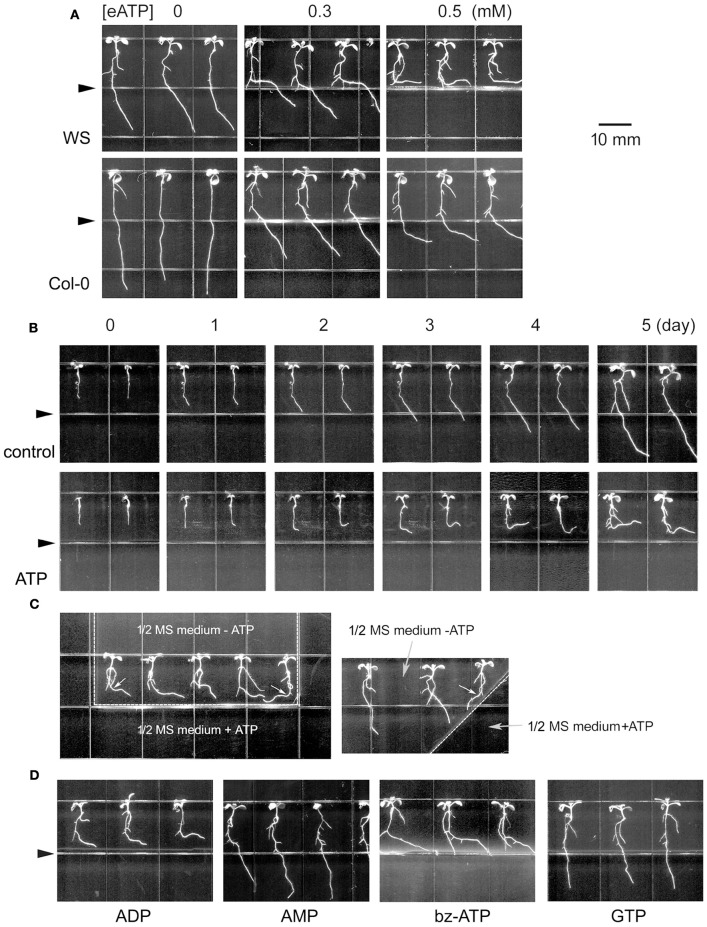
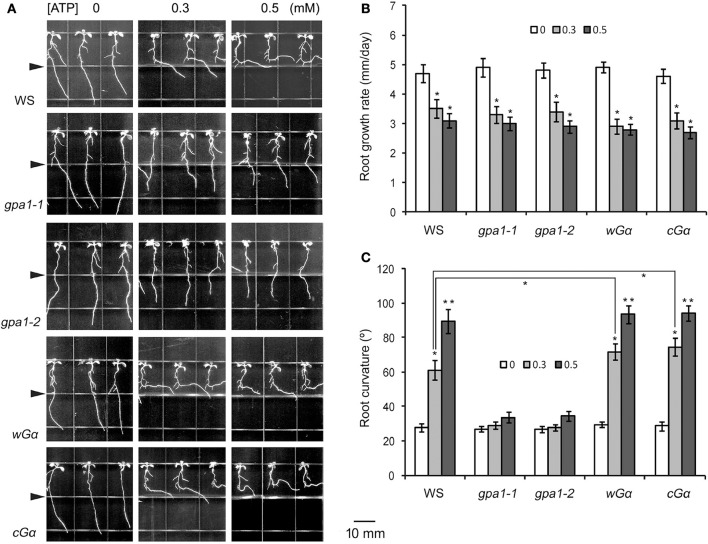
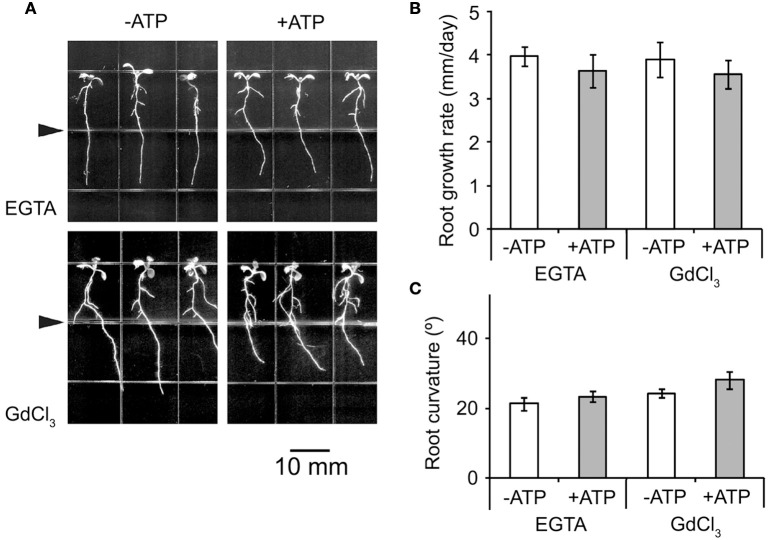
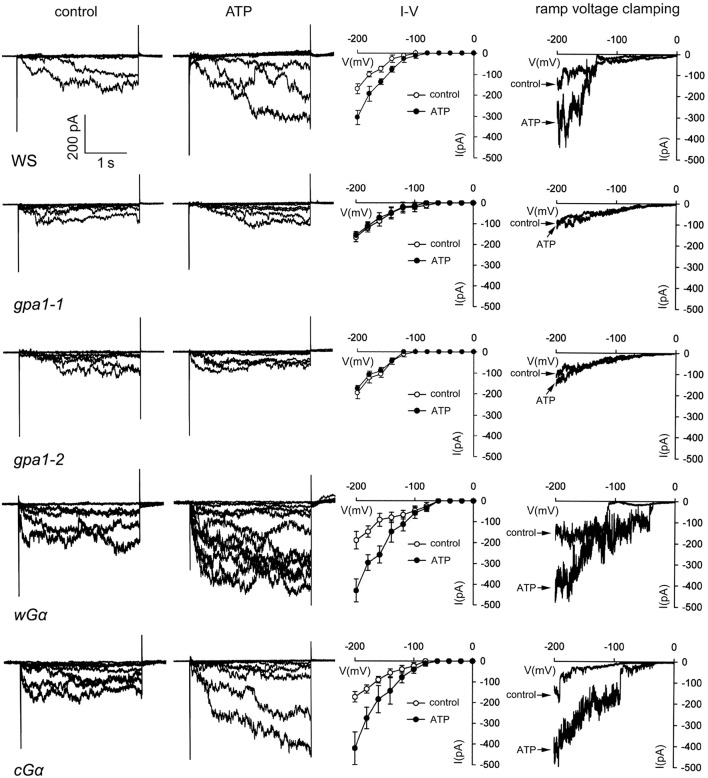
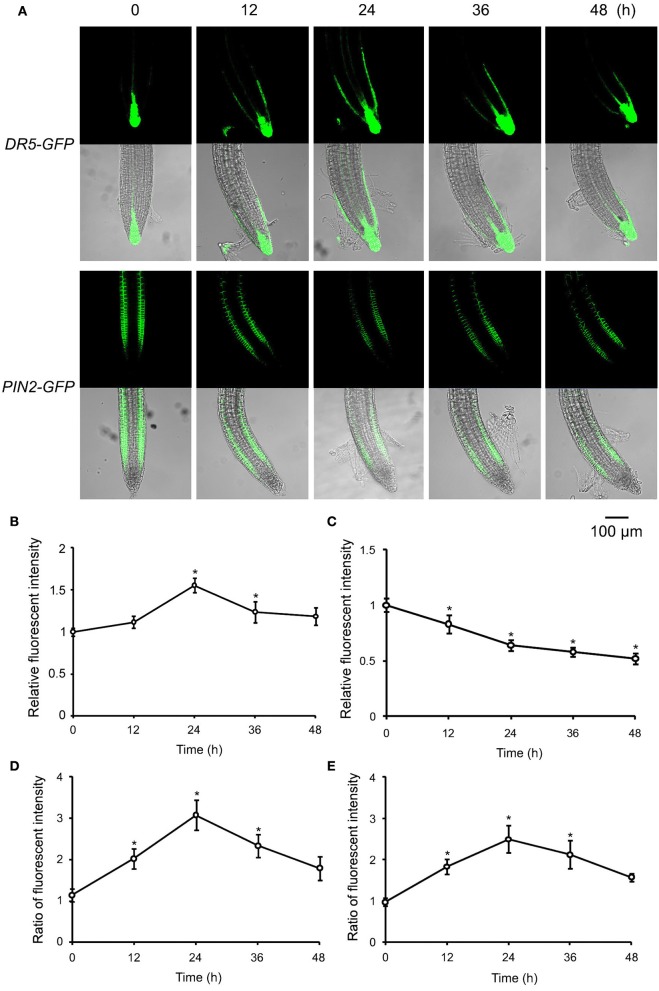
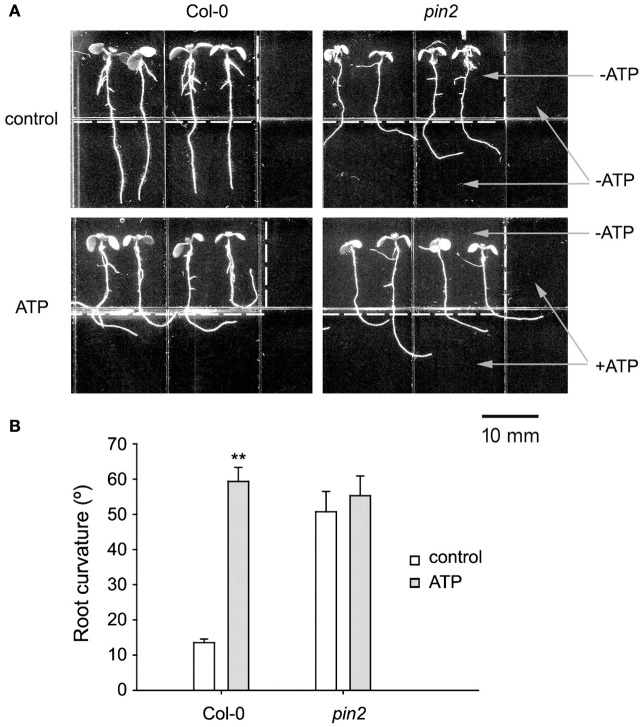
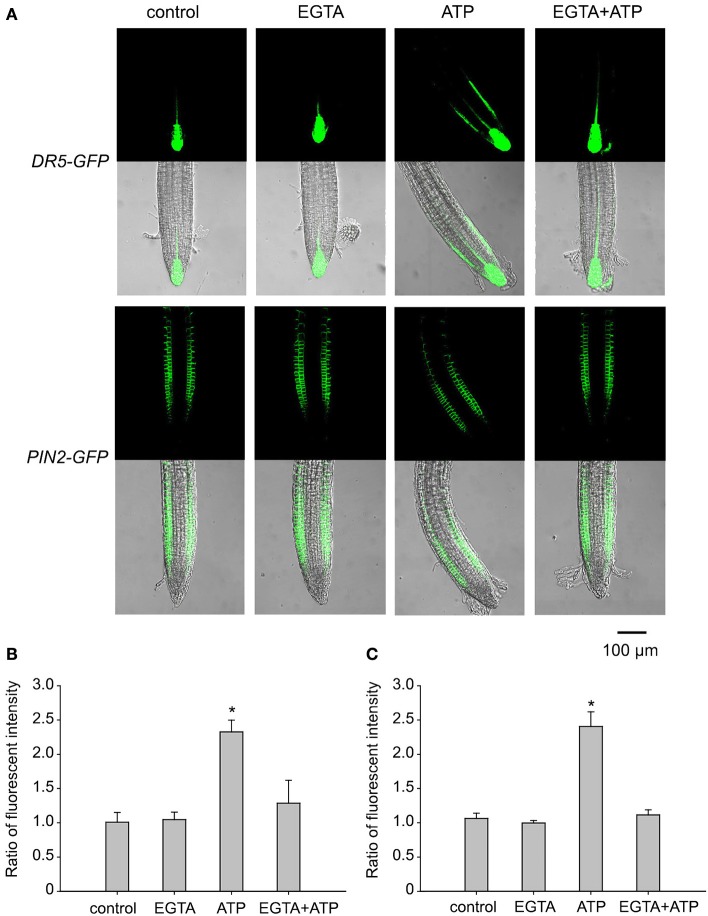
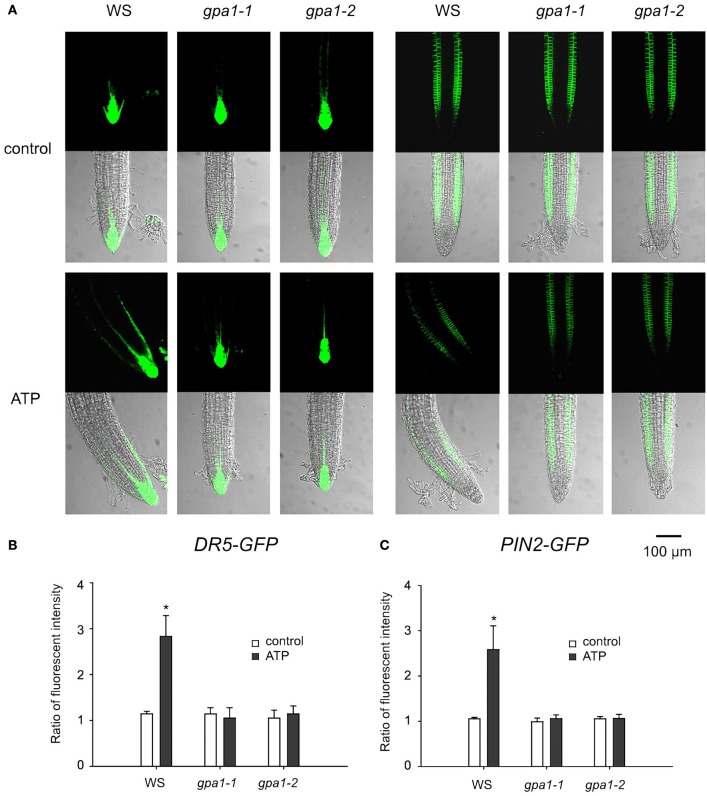
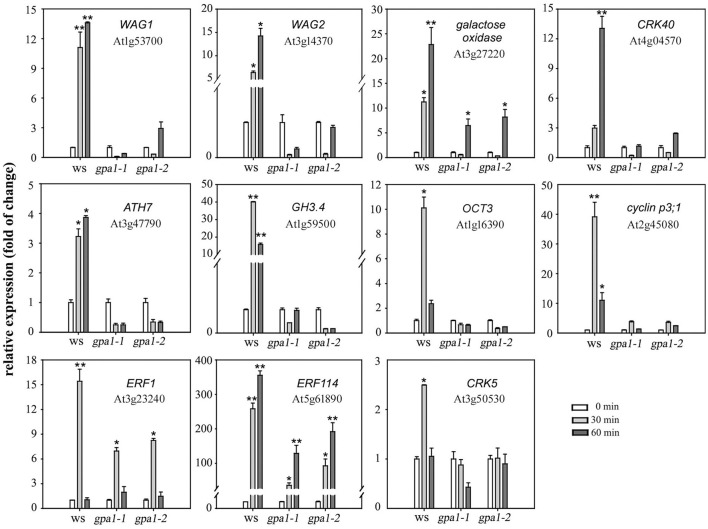
Extracellular ATP (eATP) has been reported to be involved in plant growth as a primary messenger in the apoplast. Here, roots of Arabidopsis thaliana seedlings growing in jointed medium bent upon contact with ATP-containing medium to keep away from eATP, showing a marked avoidance response. Roots responded similarly to ADP and bz-ATP but did not respond to AMP and GTP. The eATP avoidance response was reduced in loss-of-function mutants of heterotrimeric G protein α subunit (Gα) (gpa1-1 and gpa1-2) and enhanced in Gα-over-expression (OE) lines (wGα and cGα). Ethylenebis(oxyethylenenitrilo) tetraacetic acid (EGTA) and Gd3+ remarkably suppressed eATP-induced root bending. ATP-stimulated Ca2+ influx was impaired in Gα null mutants and increased in its OE lines. DR5-GFP and PIN2 were asymmetrically distributed in ATP-stimulated root tips, this effect was strongly suppressed by EGTA and diminished in Gα null mutants. In addition, some eATP-induced genes' expression was also impaired in Gα null mutants. Based on these results, we propose that heterotrimeric Gα-regulated Ca2+ influx and PIN2 distribution may be key signaling events in eATP sensing and avoidance response in Arabidopsis thaliana roots.
Keywords: Arabidopsis thaliana L.; auxin; calcium; extracellular ATP; heterotrimeric G protein.
Figures
Similar articles
-
ATANN3 Is Involved in Extracellular ATP-Regulated Auxin Distribution in Arabidopsis thaliana Seedlings.Plants (Basel). 2023 Jan 10;12(2):330. doi: 10.3390/plants12020330. Plants (Basel). 2023. PMID: 36679043 Free PMC article.
-
P2K1 Receptor, Heterotrimeric Gα Protein and CNGC2/4 Are Involved in Extracellular ATP-Promoted Ion Influx in the Pollen of Arabidopsis thaliana.Plants (Basel). 2021 Aug 23;10(8):1743. doi: 10.3390/plants10081743. Plants (Basel). 2021. PMID: 34451790 Free PMC article.
-
Extracellular ATP promotes stomatal opening of Arabidopsis thaliana through heterotrimeric G protein α subunit and reactive oxygen species.Mol Plant. 2012 Jul;5(4):852-64. doi: 10.1093/mp/ssr095. Epub 2011 Dec 2. Mol Plant. 2012. PMID: 22138967
-
Heterotrimeric G-protein participation in Arabidopsis pollen germination through modulation of a plasmamembrane hyperpolarization-activated Ca2+-permeable channel.New Phytol. 2007;176(3):550-559. doi: 10.1111/j.1469-8137.2007.02214.x. New Phytol. 2007. PMID: 17953540
-
Plant Morphology of Heterotrimeric G Protein Mutants.Plant Cell Physiol. 2016 Mar;57(3):437-45. doi: 10.1093/pcp/pcw002. Epub 2016 Jan 10. Plant Cell Physiol. 2016. PMID: 26755691 Free PMC article. Review.
Cited by
-
ATANN3 Is Involved in Extracellular ATP-Regulated Auxin Distribution in Arabidopsis thaliana Seedlings.Plants (Basel). 2023 Jan 10;12(2):330. doi: 10.3390/plants12020330. Plants (Basel). 2023. PMID: 36679043 Free PMC article.
-
Signals and Their Perception for Remodelling, Adjustment and Repair of the Plant Cell Wall.Int J Mol Sci. 2023 Apr 18;24(8):7417. doi: 10.3390/ijms24087417. Int J Mol Sci. 2023. PMID: 37108585 Free PMC article. Review.
-
Damage Signaling by Extracellular Nucleotides: A Role for Cyclic Nucleotides in Elevating Cytosolic Free Calcium?Front Plant Sci. 2021 Dec 2;12:788514. doi: 10.3389/fpls.2021.788514. eCollection 2021. Front Plant Sci. 2021. PMID: 34925428 Free PMC article.
-
P2K1 Receptor, Heterotrimeric Gα Protein and CNGC2/4 Are Involved in Extracellular ATP-Promoted Ion Influx in the Pollen of Arabidopsis thaliana.Plants (Basel). 2021 Aug 23;10(8):1743. doi: 10.3390/plants10081743. Plants (Basel). 2021. PMID: 34451790 Free PMC article.
-
Identifying Receptor Kinase Substrates Using an 8000 Peptide Kinase Client Library Enriched for Conserved Phosphorylation Sites.Mol Cell Proteomics. 2025 Mar;24(3):100926. doi: 10.1016/j.mcpro.2025.100926. Epub 2025 Feb 7. Mol Cell Proteomics. 2025. PMID: 39923935 Free PMC article.
References
-
- Balague C., Gouget A., Bouchez O., Souriac C., Haget N., Boutet-Mercey S., et al. . (2016). The Arabidopsis thaliana lectin receptor kinase LecRK-I.9 is required for full resistance to Pseudomonas syringae and affects jasmonate signalling. Mol. Plant Pathol. 18, 937–948. 10.1111/mpp.12457 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous