

Calcitox-aging counterbalanced by endogenous farnesol-like sesquiterpenoids: An undervalued evolutionarily ancient key signaling pathway
- PMID: 28919940
- PMCID: PMC5595427
- DOI: 10.1080/19420889.2017.1341024
Calcitox-aging counterbalanced by endogenous farnesol-like sesquiterpenoids: An undervalued evolutionarily ancient key signaling pathway
Abstract
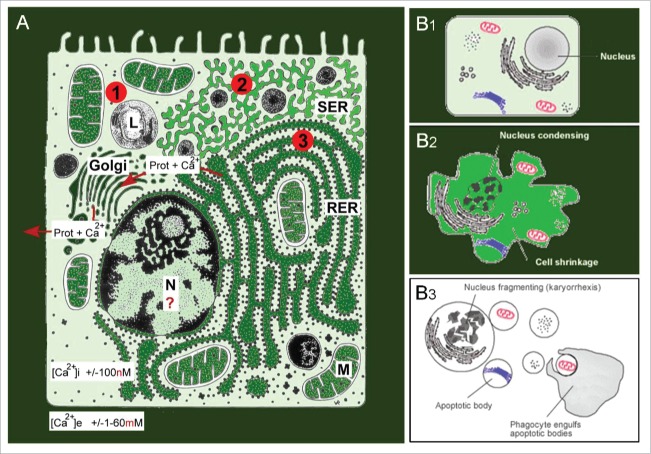
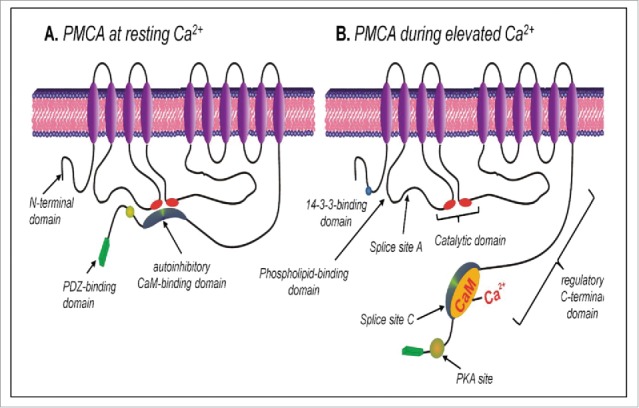
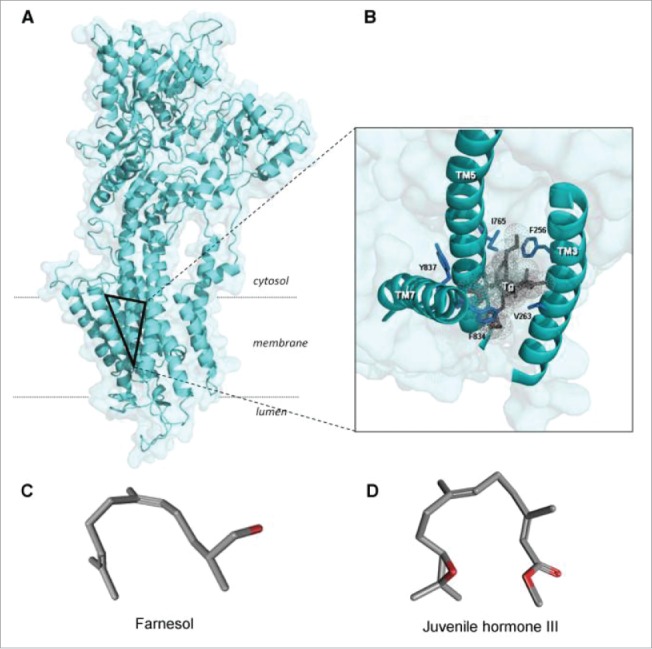
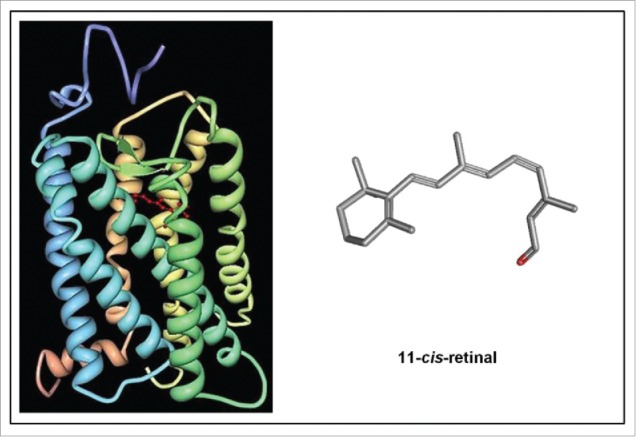
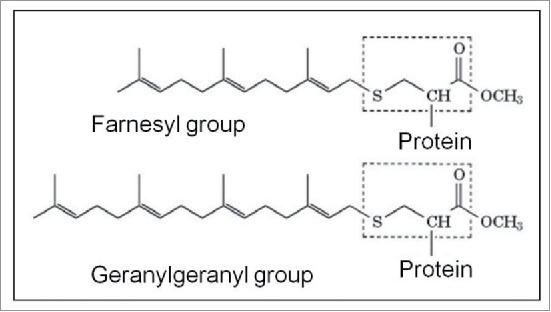
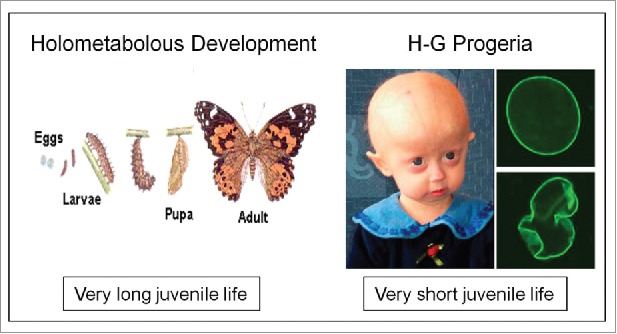
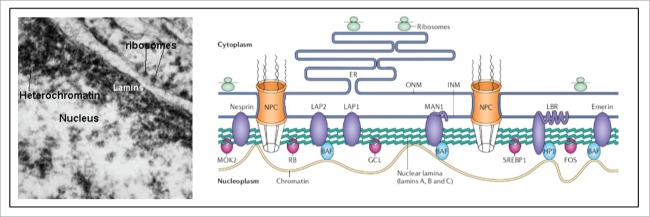
Cells are powerful miniature electrophoresis chambers, at least during part of their life cycle. They die at the moment the voltage gradient over their plasma membrane, and their ability to drive a self-generated electric current carried by inorganic ions through themselves irreversibly collapses. Senescence is likely due to the progressive, multifactorial damage to the cell's electrical system. This is the essence of the "Fading electricity theory of aging" (De Loof et al., Aging Res. Rev. 2013;12:58-66). "Biologic electric current" is not carried by electrons, but by inorganic ions. The major ones are H+, Na+, K+, Ca2+, Mg2+, Cl- and HCO3-. Ca2+ and H+ in particular are toxic to cells. At rising concentrations, they can alter the 3D-conformation of chromatin and some (e.g. cytoskeletal) proteins: Calcitox and Protontox. This paper only focuses on Calcitox and endogenous sesquiterpenoids. pH-control and Ca2+-homeostasis have been shaped to near perfection during billions of years of evolution. The role of Ca2+ in some aspects of aging, e.g., as causal to neurodegenerative diseases is still debated. The main anti-Calcitox mechanism is to keep free cytoplasmic Ca2+ as low as possible. This can be achieved by restricting the passive influx of Ca2+ through channels in the plasma membrane, and by maximizing the active extrusion of excess Ca2+ e.g., by means of different types of Ca2+-ATPases. Like there are mechanisms that antagonize the toxic effects of Reactive Oxygen Species (ROS), there must also exist endogenous tools to counteract Calcitox. During a re-evaluation of which mechanism(s) exactly initiates the fast aging that accompanies induction of metamorphosis in insects, a causal relationship between absence of an endogenous sesquiterpenoid, namely the farnesol ester named "juvenile hormone," and disturbed Ca2+-homeostasis was suggested. In this paper, this line of thinking is further explored and extended to vertebrate physiology. A novel concept emerges: horseshoe-shaped sesquiterpenoids seem to act as "inbrome" agonists with the function of a "chemical valve" or "spring" in some types of multi-helix transmembrane proteins (intramolecular prenylation), from bacterial rhodopsins to some types of GPCRs and ion pumps, in particular the SERCA-Ca2+-pump. This further underpins the Fading Electricity Theory of Aging.
Keywords: Alzheimer; SERCA; cytoskeleton; electrome; farnesol; juvenile hormone; prenylation; retinal; senescence; thapsigargin.
Figures




References
-
- Cannon ML. What is aging? Disease-a-Month 2015; 61:454-9; PMID:26519984; https://doi.org/10.1016/j.disamonth.2015.09.002 - DOI - PubMed
-
- MS, King M. Biological theories of aging. Disease-a-Month 2015; 61:460-66; PMID:26490576; https://doi.org/10.1016/j.disamonth.2015.09.005 - DOI - PubMed
-
- Pinto da Costa J, Vitorino R, Silva GM, Vogel C, Duarte AC, Rocha-Santos T. A synopsis on aging – Theories, mechanisms and future prospects. Ageing Res Rev 2016; 29:90-112; PMID:27353257; https://doi.org/10.1016/j.arr.2016.06.005 - DOI - PMC - PubMed
-
- Adam G, Perrimon N, Noselli S. The retinoic-like juvenile hormone controls the looping of left-right asymmetric organs in Drosophila. Development 2003; 130:2397-06; PMID:12702654; https://doi.org/10.1242/dev.00460 - DOI - PubMed
-
- Palczewski K. Chemistry and biology of vision. J Biol Chem 2013; 287:1612-9; https://doi.org/10.1074/jbc.R111.301150 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous