

Human tumor-derived exosomes (TEX) regulate Treg functions via cell surface signaling rather than uptake mechanisms
- PMID: 28919985
- PMCID: PMC5593709
- DOI: 10.1080/2162402X.2016.1261243
Human tumor-derived exosomes (TEX) regulate Treg functions via cell surface signaling rather than uptake mechanisms
Abstract
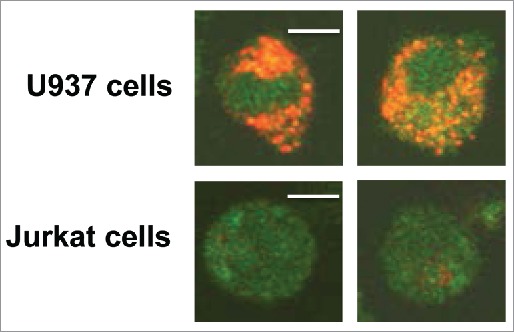
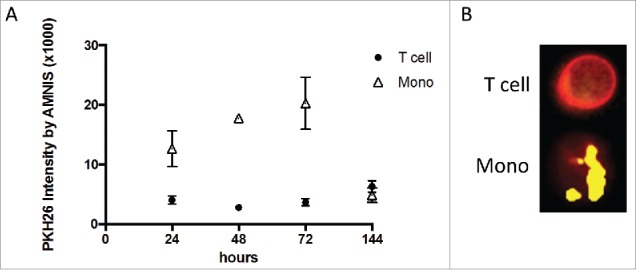
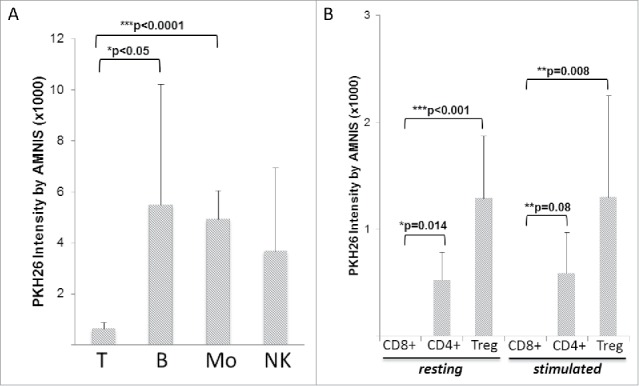
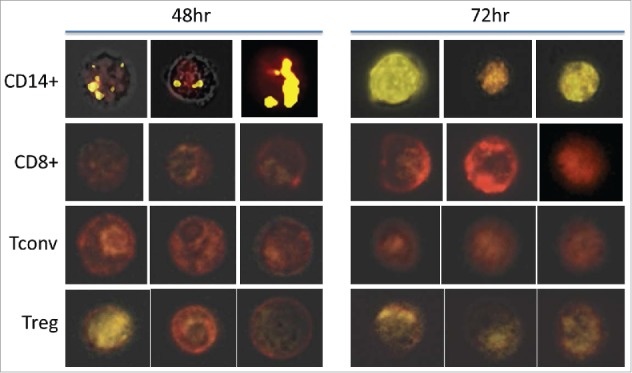
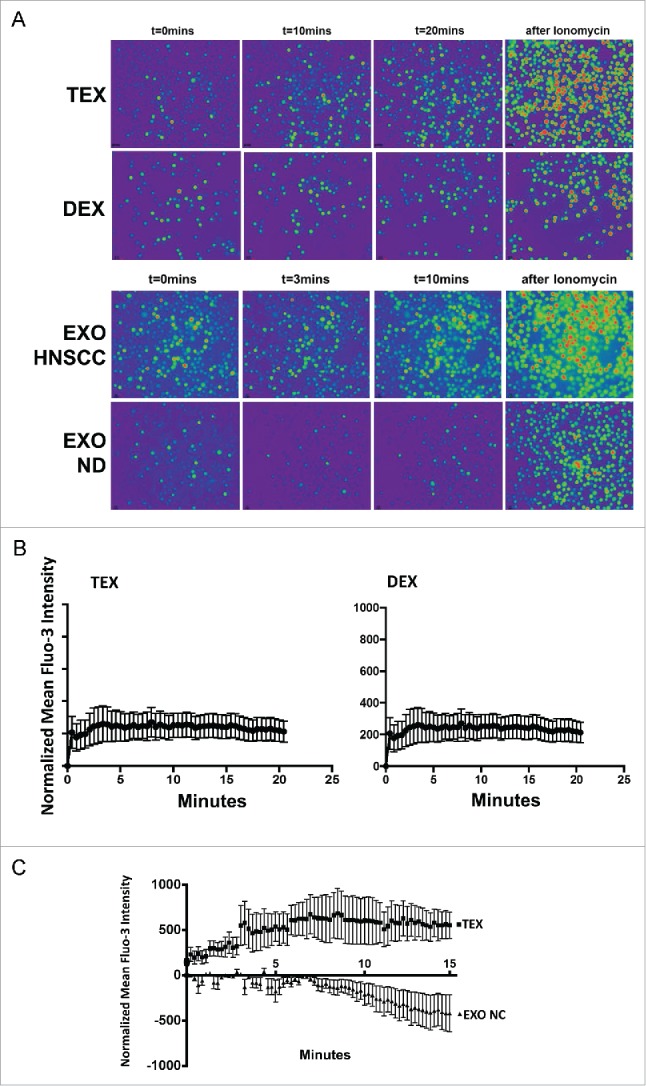
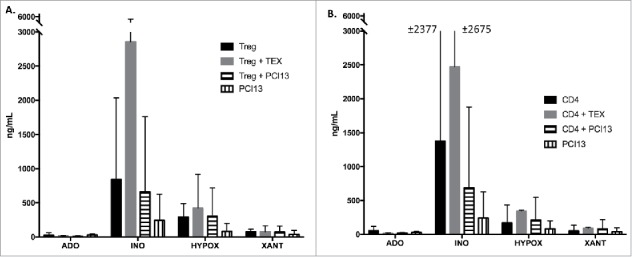
Tumor-derived exosomes (TEX) are ubiquitously present in the tumor microenvironment and plasma of cancer patients. TEX carry a cargo of multiple stimulatory and inhibitory molecules and deliver them to recipient cells, serving as a communication network for the tumor. The mechanisms TEX use for delivering messages to recipient cells were evaluated using PKH26-labeled TEX produced by cultured human tumor cells, exosomes produced by dendritic cells-derived exosomes (DEX), or exosomes isolated from plasma of cancer patients (EXO). Human T-cell subsets, B cells, NK cells, and monocytes were co-incubated with TEX, DEX, or EXO and binding or internalization of labeled vesicles was evaluated by confocal microscopy and/or Amnis-based flow cytometry. Vesicle-induced Ca2+ influx in recipient T cells was monitored, and TEX-induced inosine production in Treg was determined by mass spectrometry. In contrast to B cells, NK cells or monocytes, conventional T cells did not internalize labeled vesicles. Minimal exosome uptake was only evident in Treg following prolonged co-incubation with TEX. All exosomes induced Ca2+ influx in T cells, with TEX and EXO isolated from cancer patients' plasma delivering the strongest, sustained signaling to Treg. Such sustained signaling resulted in the significant upregulation of the conversion of extracellular ATP to inosine (adenosine metabolite) by Treg, suggesting that TEX signaling could have functional consequences in these recipient cells. Thus, modulation of Treg suppressor functions by TEX is mediated by mechanisms dependent on cell surface signaling and does not require TEX internalization by recipient cells.
Keywords: Ca2+ flux; T cell subsets; exosome uptake; regulatory T cells; tumor-derived exosomes.
Figures
Similar articles
-
Tumor-derived exosomes regulate expression of immune function-related genes in human T cell subsets.Sci Rep. 2016 Feb 4;6:20254. doi: 10.1038/srep20254. Sci Rep. 2016. PMID: 26842680 Free PMC article.
-
Immune Suppressive Effects of Plasma-Derived Exosome Populations in Head and Neck Cancer.Cancers (Basel). 2020 Jul 21;12(7):1997. doi: 10.3390/cancers12071997. Cancers (Basel). 2020. PMID: 32708274 Free PMC article.
-
Circulating exosomes measure responses to therapy in head and neck cancer patients treated with cetuximab, ipilimumab, and IMRT.Oncoimmunology. 2019 Apr 24;8(7):1593805. doi: 10.1080/2162402X.2019.1593805. eCollection 2019. Oncoimmunology. 2019. PMID: 31143513 Free PMC article.
-
Exosome and mesenchymal stem cell cross-talk in the tumor microenvironment.Semin Immunol. 2018 Feb;35:69-79. doi: 10.1016/j.smim.2017.12.003. Epub 2017 Dec 27. Semin Immunol. 2018. PMID: 29289420 Free PMC article. Review.
-
Tumor-Derived Exosomes (TEX) and Their Role in Immuno-Oncology.Int J Mol Sci. 2021 Jun 9;22(12):6234. doi: 10.3390/ijms22126234. Int J Mol Sci. 2021. PMID: 34207762 Free PMC article. Review.
Cited by
-
Trial watch: dendritic cell vaccination for cancer immunotherapy.Oncoimmunology. 2019 Jul 18;8(11):e1638212. doi: 10.1080/2162402X.2019.1638212. eCollection 2019. Oncoimmunology. 2019. PMID: 31646087 Free PMC article. Review.
-
Tumor-derived extracellular vesicles regulate tumor-infiltrating regulatory T cells via the inhibitory immunoreceptor CD300a.Elife. 2021 Nov 9;10:e61999. doi: 10.7554/eLife.61999. Elife. 2021. PMID: 34751648 Free PMC article.
-
Toll-like Receptor-4 Activation Boosts the Immunosuppressive Properties of Tumor Cells-derived Exosomes.Sci Rep. 2019 Jun 11;9(1):8457. doi: 10.1038/s41598-019-44949-y. Sci Rep. 2019. PMID: 31186484 Free PMC article.
-
Deconstructing Pancreatic Cancer Using Next Generation-Omic Technologies-From Discovery to Knowledge-Guided Platforms for Better Patient Management.Front Cell Dev Biol. 2022 Jan 13;9:795735. doi: 10.3389/fcell.2021.795735. eCollection 2021. Front Cell Dev Biol. 2022. PMID: 35096825 Free PMC article. Review.
-
Tumor-Derived Extracellular Vesicles: Their Role in Immune Cells and Immunotherapy.Int J Nanomedicine. 2021 Aug 10;16:5395-5409. doi: 10.2147/IJN.S313912. eCollection 2021. Int J Nanomedicine. 2021. PMID: 34408415 Free PMC article. Review.
References
-
- Raposo G, Stoorvogel W. Extracellular vesicles: exosomes, microvesicles, and friends. J Cell Biol 2013; 200:373-83; PMID:23420871; https://doi.org/10.1083/jcb.201211138 - DOI - PMC - PubMed
-
- Figueiro F, Muller L, Funk S, Jackson EK, Battastini AM, Whiteside TL. Phenotypic and functional characteristics of CD39high human regulatory B cells (Breg). Oncoimmunology 2016; 5:e1082703; PMID:27057473; https://doi.org/10.1080/2162402X.2015.1082703 - DOI - PMC - PubMed
-
- Keller S, Ridinger J, Rupp AK, Janssen JW, Altevogt P. Body fluid derived exosomes as a novel template for clinical diagnostics. J Transl Med 2011; 9:86; PMID:21651777; https://doi.org/10.1186/1479-5876-9-86 - DOI - PMC - PubMed
-
- Brinton LT, Sloane HS, Kester M, Kelly KA. Formation and role of exosomes in cancer. Cell Mol Life Sci 2015; 72:659-71; PMID:25336151; https://doi.org/10.1007/s00018-014-1764-3 - DOI - PMC - PubMed
-
- Kowal J, Tkach M, Thery C. Biogenesis and secretion of exosomes. Curr Opin Cell Biol 2014; 29:116-25; PMID:24959705; https://doi.org/10.1016/j.ceb.2014.05.004 - DOI - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous