

The N6-methyladenosine (m6A)-forming enzyme METTL3 controls myeloid differentiation of normal hematopoietic and leukemia cells
- PMID: 28920958
- PMCID: PMC5677536
- DOI: 10.1038/nm.4416
The N6-methyladenosine (m6A)-forming enzyme METTL3 controls myeloid differentiation of normal hematopoietic and leukemia cells
Abstract
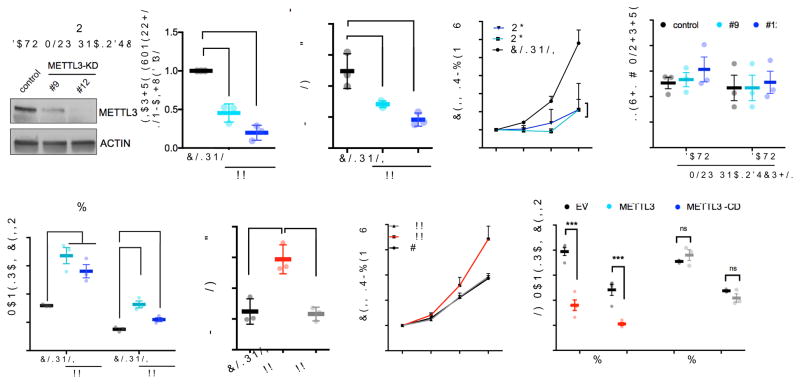
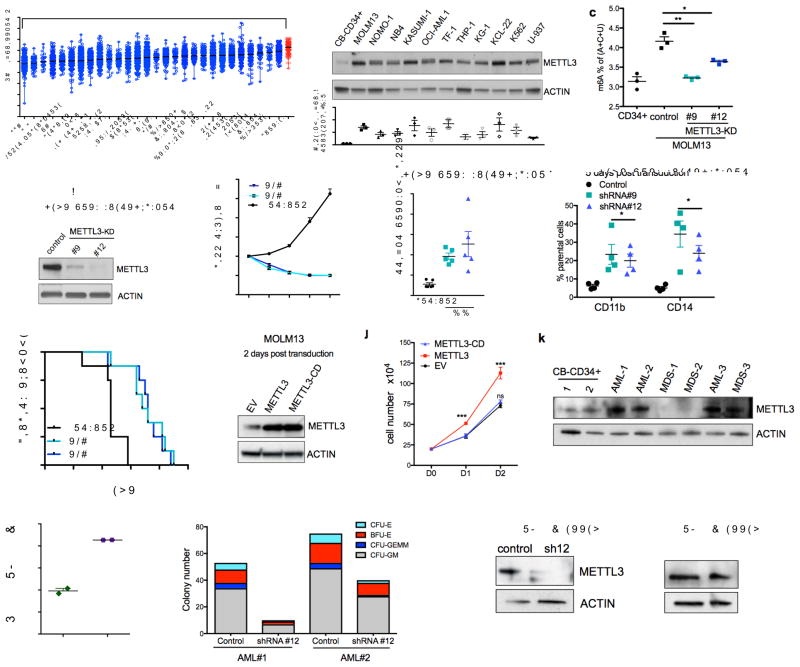
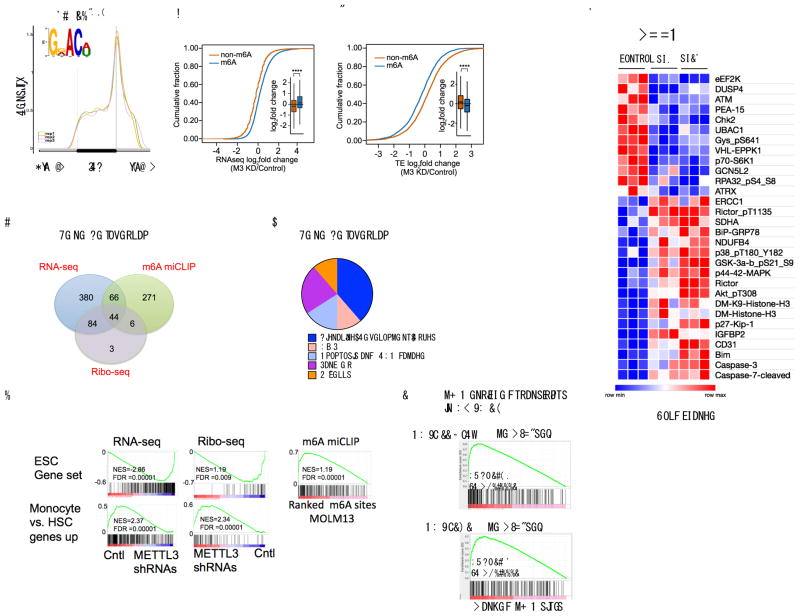
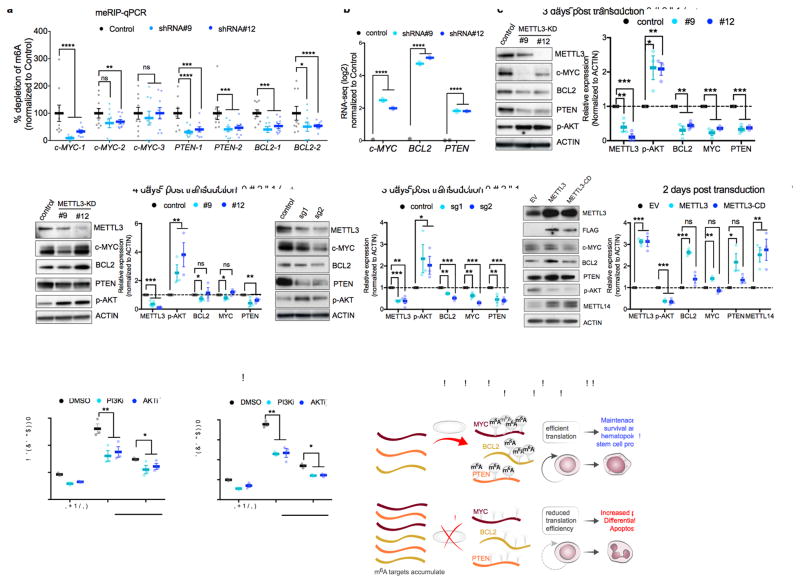
N6-methyladenosine (m6A) is an abundant nucleotide modification in mRNA that is required for the differentiation of mouse embryonic stem cells. However, it remains unknown whether the m6A modification controls the differentiation of normal and/or malignant myeloid hematopoietic cells. Here we show that shRNA-mediated depletion of the m6A-forming enzyme METTL3 in human hematopoietic stem/progenitor cells (HSPCs) promotes cell differentiation, coupled with reduced cell proliferation. Conversely, overexpression of wild-type METTL3, but not of a catalytically inactive form of METTL3, inhibits cell differentiation and increases cell growth. METTL3 mRNA and protein are expressed more abundantly in acute myeloid leukemia (AML) cells than in healthy HSPCs or other types of tumor cells. Furthermore, METTL3 depletion in human myeloid leukemia cell lines induces cell differentiation and apoptosis and delays leukemia progression in recipient mice in vivo. Single-nucleotide-resolution mapping of m6A coupled with ribosome profiling reveals that m6A promotes the translation of c-MYC, BCL2 and PTEN mRNAs in the human acute myeloid leukemia MOLM-13 cell line. Moreover, loss of METTL3 leads to increased levels of phosphorylated AKT, which contributes to the differentiation-promoting effects of METTL3 depletion. Overall, these results provide a rationale for the therapeutic targeting of METTL3 in myeloid leukemia.
Conflict of interest statement
There is no competing interest.
Figures
Comment in
-
An epitranscriptomic vulnerability in myeloid malignancies.Nat Med. 2017 Nov 7;23(11):1252-1254. doi: 10.1038/nm.4435. Nat Med. 2017. PMID: 29117175 No abstract available.
-
Meddling with METTLs in Normal and Leukemia Stem Cells.Cell Stem Cell. 2018 Feb 1;22(2):139-141. doi: 10.1016/j.stem.2018.01.013. Cell Stem Cell. 2018. PMID: 29395048
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
