

Bone marrow type 2 innate lymphoid cells: a local source of interleukin-5 in interleukin-33-driven eosinophilia
- PMID: 28921511
- PMCID: PMC5765380
- DOI: 10.1111/imm.12842
Bone marrow type 2 innate lymphoid cells: a local source of interleukin-5 in interleukin-33-driven eosinophilia
Abstract
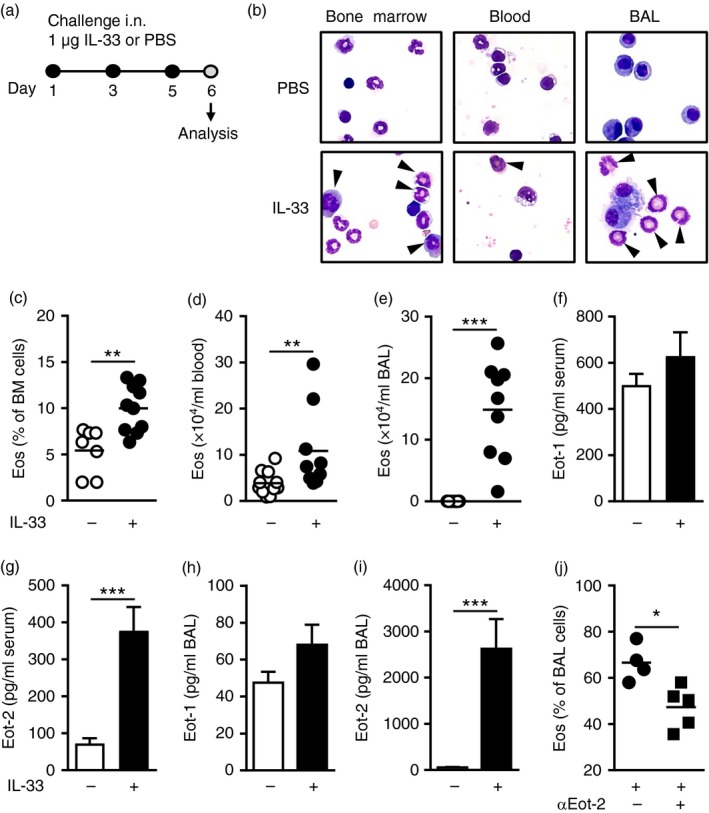
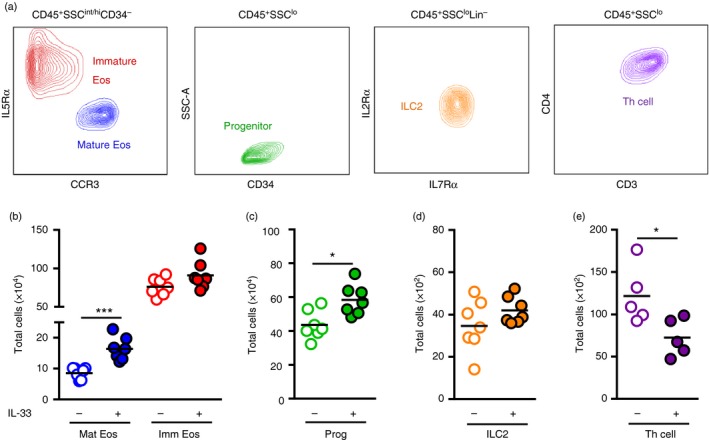
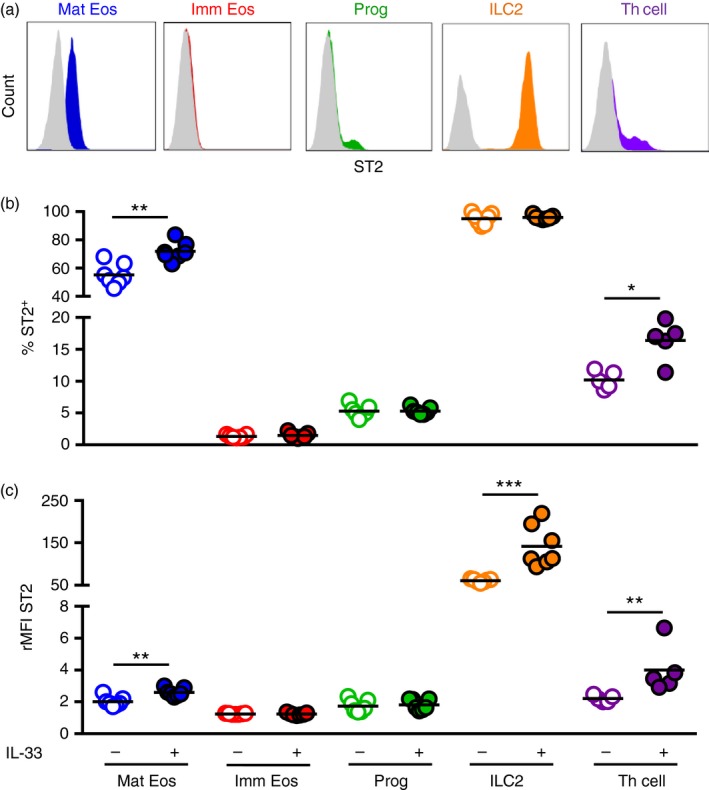
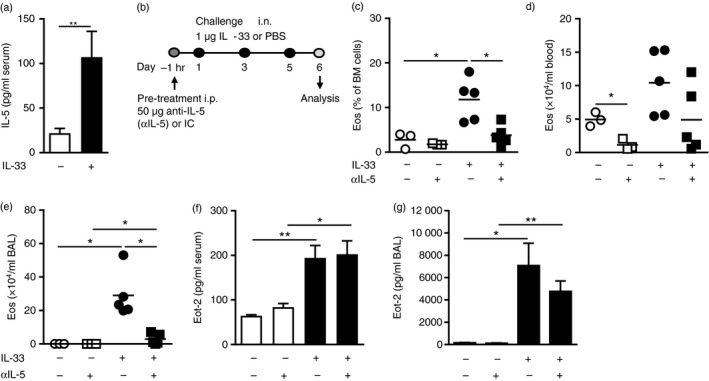
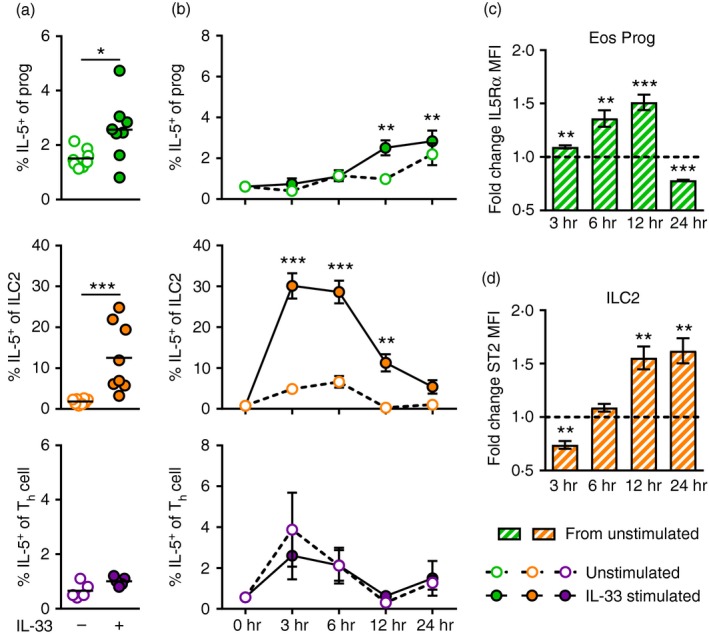
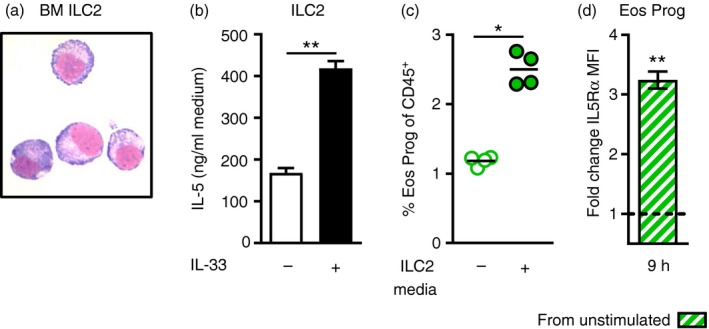
T helper type 2 (Th2) cells, type 2 innate lymphoid cells (ILC2s) and eosinophil progenitors have previously been described to produce interleukin-5 (IL-5) in the airways upon allergen provocation or by direct administration of IL-33. Eosinophilic airway inflammation is known to be associated with IL-5-dependent eosinophil development in the bone marrow, however, the source of IL-5 remains unclear. T helper cells, ILC2s and CD34+ progenitors have been proposed to be involved in this process, therefore, we investigated whether these cells are taking part in eosinophilopoiesis by producing IL-5 locally in the bone marrow in IL-33-driven inflammation. Airway exposure with IL-33 led to eosinophil infiltration in airways and elevated eotaxin-2/CCL24. Importantly, IL-5 production as well as expression of the IL-33 receptor increased in ILC2s in the bone marrow under this treatment. A small but significant induction of IL-5 was also found in CD34+ progenitors but not in T helper cells. Similar results were obtained by in vitro stimulation with IL-33 where ILC2s rapidly produced large amounts of IL-5, which coincided with the induction of eosinophil hematopoiesis. IL-33-mediated eosinophil production was indeed dependent on IL-5 as both airway and bone marrow eosinophils decreased in mice treated with anti-IL-5 in combination with IL-33. Interestingly, the responsiveness of ILC2s to IL-33 as well as IL-33-induced eotaxin-2/CCL24 were independent of the levels of IL-5. In summary, we demonstrate for the first time that IL-33 acts directly on bone marrow ILC2s, making them an early source of IL-5 and part of a process that is central in IL-33-driven eosinophilia.
Keywords: IL-33; IL-5; ILC2; bone marrow; eosinophilia.
© 2017 John Wiley & Sons Ltd.
Figures
Similar articles
-
Interplay Between the IL-33/ST2 Axis and Bone Marrow ILC2s in Protease Allergen-Induced IL-5-Dependent Eosinophilia.Front Immunol. 2020 Jun 2;11:1058. doi: 10.3389/fimmu.2020.01058. eCollection 2020. Front Immunol. 2020. PMID: 32582171 Free PMC article.
-
House Dust Mite Induces Bone Marrow IL-33-Responsive ILC2s and TH Cells.Int J Mol Sci. 2020 May 26;21(11):3751. doi: 10.3390/ijms21113751. Int J Mol Sci. 2020. PMID: 32466530 Free PMC article.
-
IL-33 mediates reactive eosinophilopoiesis in response to airborne allergen exposure.Allergy. 2016 Jul;71(7):977-88. doi: 10.1111/all.12861. Epub 2016 Mar 2. Allergy. 2016. PMID: 26864308 Free PMC article.
-
Haemopoietic processes in allergic disease: eosinophil/basophil development.Clin Exp Allergy. 2009 Sep;39(9):1297-306. doi: 10.1111/j.1365-2222.2009.03325.x. Epub 2009 Jul 20. Clin Exp Allergy. 2009. PMID: 19622087 Review.
-
Type 2 innate lymphoid cells: at the cross-roads in allergic asthma.Semin Immunopathol. 2016 Jul;38(4):483-96. doi: 10.1007/s00281-016-0556-2. Epub 2016 Mar 10. Semin Immunopathol. 2016. PMID: 26965110 Free PMC article. Review.
Cited by
-
Tissue-Specific Molecular Markers and Heterogeneity in Type 2 Innate Lymphoid Cells.Front Immunol. 2021 Oct 25;12:757967. doi: 10.3389/fimmu.2021.757967. eCollection 2021. Front Immunol. 2021. PMID: 34759931 Free PMC article. Review.
-
Rapamycin Dampens Inflammatory Properties of Bone Marrow ILC2s in IL-33-Induced Eosinophilic Airway Inflammation.Front Immunol. 2022 Jun 3;13:915906. doi: 10.3389/fimmu.2022.915906. eCollection 2022. Front Immunol. 2022. PMID: 35720347 Free PMC article.
-
Molecular Pathogenesis and Treatment Perspectives for Hypereosinophilia and Hypereosinophilic Syndromes.Int J Mol Sci. 2021 Jan 6;22(2):486. doi: 10.3390/ijms22020486. Int J Mol Sci. 2021. PMID: 33418988 Free PMC article. Review.
-
IL-33/ST2 signaling in ILC2s drives exhaustion and myeloid skewing of HSCs in response to hematopoietic stress and aging.iScience. 2025 Apr 8;28(5):112378. doi: 10.1016/j.isci.2025.112378. eCollection 2025 May 16. iScience. 2025. PMID: 40384929 Free PMC article.
-
Eosinophilic inflammation: An Appealing Target for Pharmacologic Treatments in Severe Asthma.Biomedicines. 2022 Sep 3;10(9):2181. doi: 10.3390/biomedicines10092181. Biomedicines. 2022. PMID: 36140282 Free PMC article. Review.
References
-
- Wardlaw AJ, Brightling C, Green R, Woltmann G, Pavord I. Eosinophils in asthma and other allergic diseases. Br Med Bull 2000; 56:985–1003. - PubMed
-
- Rothenberg ME, Hogan SP. The eosinophil. Annu Rev Immunol 2006; 24:147–74. - PubMed
-
- Ema H, Suda T, Nagayoshi K, Miura Y, Civin CI, Nakauchi H. Target cells for granulocyte colony‐stimulating factor, interleukin‐3, and interleukin‐5 in differentiation pathways of neutrophils and eosinophils. Blood 1990; 76:1956–61. - PubMed
-
- Yang M, Hogan SP, Mahalingam S, Pope SM, Zimmermann N, Fulkerson P et al Eotaxin‐2 and IL‐5 cooperate in the lung to regulate IL‐13 production and airway eosinophilia and hyperreactivity. J Allergy Clin Immunol 2003; 112:935–43. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
