

α-SNAP is expressed in mouse ovarian granulosa cells and plays a key role in folliculogenesis and female fertility
- PMID: 28924180
- PMCID: PMC5603506
- DOI: 10.1038/s41598-017-12292-9
α-SNAP is expressed in mouse ovarian granulosa cells and plays a key role in folliculogenesis and female fertility
Erratum in
-
Publisher Correction: α-SNAP is expressed in mouse ovarian granulosa cells and plays a key role in folliculogenesis and female fertility.Sci Rep. 2020 Mar 25;10(1):5691. doi: 10.1038/s41598-020-61863-w. Sci Rep. 2020. PMID: 32214131 Free PMC article.
Abstract
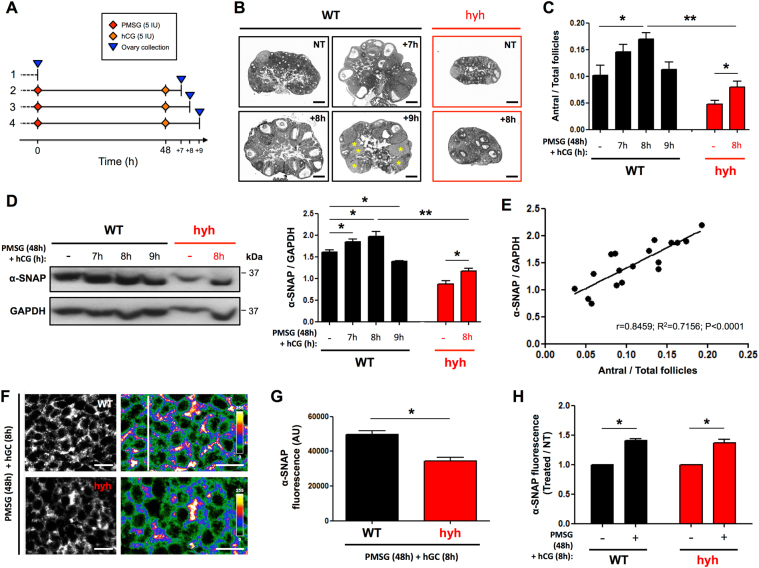
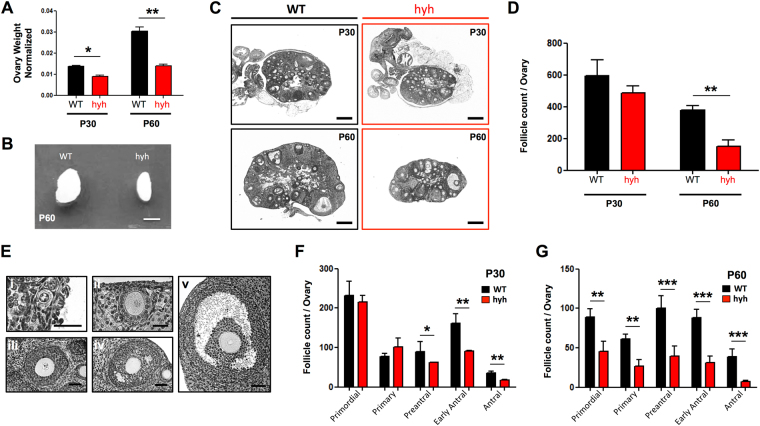
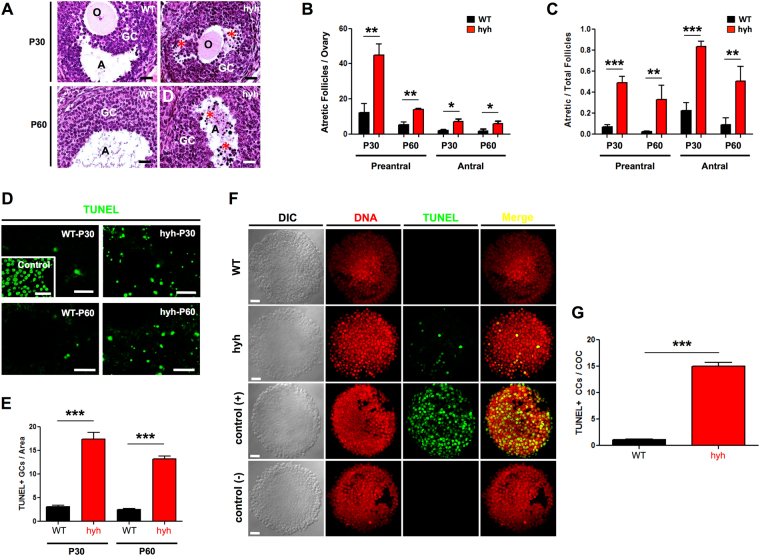
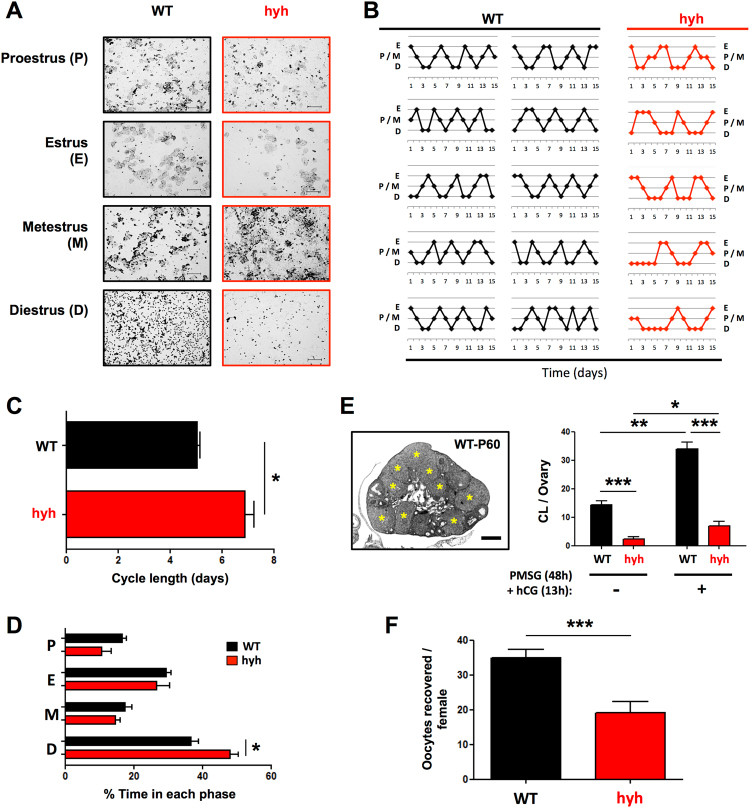
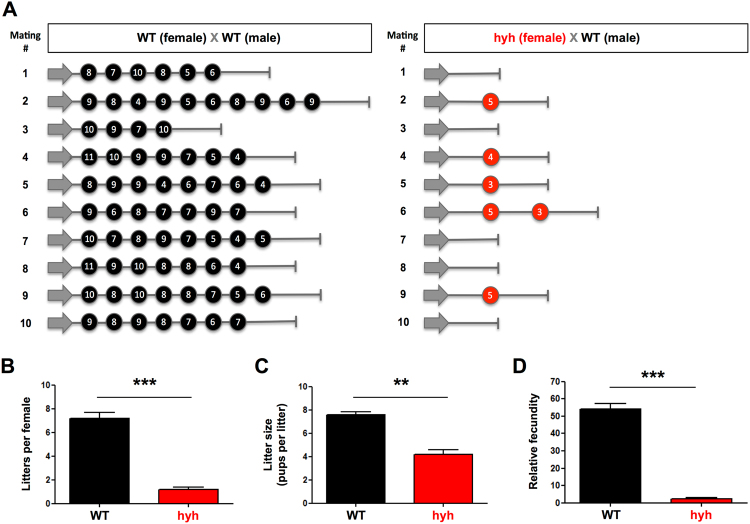
The balance between ovarian folliculogenesis and follicular atresia is critical for female fertility and is strictly regulated by a complex network of neuroendocrine and intra-ovarian signals. Despite the numerous functions executed by granulosa cells (GCs) in ovarian physiology, the role of multifunctional proteins able to simultaneously coordinate/modulate several cellular pathways is unclear. Soluble N-ethylmaleimide-sensitive factor (NSF) attachment protein (α-SNAP) is a multifunctional protein that participates in SNARE-mediated membrane fusion events. In addition, it regulates cell-to-cell adhesion, AMPK signaling, autophagy and apoptosis in different cell types. In this study we examined the expression pattern of α-SNAP in ovarian tissue and the consequences of α-SNAP (M105I) mutation (hyh mutation) in folliculogenesis and female fertility. Our results showed that α-SNAP protein is highly expressed in GCs and its expression is modulated by gonadotropin stimuli. On the other hand, α-SNAP-mutant mice show a reduction in α-SNAP protein levels. Moreover, increased apoptosis of GCs and follicular atresia, reduced ovulation rate, and a dramatic decline in fertility is observed in α-SNAP-mutant females. In conclusion, α-SNAP plays a critical role in the balance between follicular development and atresia. Consequently, a reduction in its expression/function (M105I mutation) causes early depletion of ovarian follicles and female subfertility.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures

Similar articles
-
Hyal-1 but not Hyal-3 deficiency has an impact on ovarian folliculogenesis and female fertility by altering the follistatin/activin/Smad3 pathway and the apoptotic process.J Cell Physiol. 2012 May;227(5):1911-22. doi: 10.1002/jcp.22919. J Cell Physiol. 2012. PMID: 21732362
-
Alterations in gonadotropin, apoptotic and metabolic pathways in granulosa cells warrant superior fertility of the Dummerstorf high fertility mouse line 1.J Ovarian Res. 2023 Feb 4;16(1):32. doi: 10.1186/s13048-023-01113-5. J Ovarian Res. 2023. PMID: 36739419 Free PMC article.
-
Pleiotropic effects of alpha-SNAP M105I mutation on oocyte biology: ultrastructural and cellular changes that adversely affect female fertility in mice.Sci Rep. 2019 Nov 22;9(1):17374. doi: 10.1038/s41598-019-53574-8. Sci Rep. 2019. PMID: 31758001 Free PMC article.
-
Regulation of cell death and cell survival gene expression during ovarian follicular development and atresia.Front Biosci. 2003 Jan 1;8:d222-37. doi: 10.2741/949. Front Biosci. 2003. PMID: 12456353 Review.
-
MicroRNAs: New Insight in Modulating Follicular Atresia: A Review.Int J Mol Sci. 2017 Feb 9;18(2):333. doi: 10.3390/ijms18020333. Int J Mol Sci. 2017. PMID: 28208755 Free PMC article. Review.
Cited by
-
Effect of Disulfiram on the Reproductive Capacity of Female Mice.Int J Mol Sci. 2023 Jan 25;24(3):2371. doi: 10.3390/ijms24032371. Int J Mol Sci. 2023. PMID: 36768698 Free PMC article.
-
Zika virus dysregulates human Sertoli cell proteins involved in spermatogenesis with little effect on tight junctions.PLoS Negl Trop Dis. 2020 Jun 8;14(6):e0008335. doi: 10.1371/journal.pntd.0008335. eCollection 2020 Jun. PLoS Negl Trop Dis. 2020. PMID: 32511241 Free PMC article.
-
Alpha-SNAP (M105I) mutation promotes neuronal differentiation of neural stem/progenitor cells through overactivation of AMPK.Front Cell Dev Biol. 2023 Apr 11;11:1061777. doi: 10.3389/fcell.2023.1061777. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 37113766 Free PMC article.
-
Impact of NAD+ metabolism on ovarian aging.Immun Ageing. 2023 Dec 2;20(1):70. doi: 10.1186/s12979-023-00398-w. Immun Ageing. 2023. PMID: 38041117 Free PMC article. Review.
-
In vitro evaluation of exocytosis-associated SNARE molecules in human granulosa cells in polycystic ovary syndrome.J Assist Reprod Genet. 2024 Jan;41(1):49-61. doi: 10.1007/s10815-023-02967-w. Epub 2023 Nov 23. J Assist Reprod Genet. 2024. PMID: 37993579 Free PMC article.
References
-
- Schwartz, N. B. In Neuroendocrinology in Physiology and Medicine (ed P. M. Conn, Freeman ME) 135–146 (Humana Press, 2000).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
