

PDK4 Inhibits Cardiac Pyruvate Oxidation in Late Pregnancy
- PMID: 28928113
- PMCID: PMC5722682
- DOI: 10.1161/CIRCRESAHA.117.311456
PDK4 Inhibits Cardiac Pyruvate Oxidation in Late Pregnancy
Abstract
Rationale: Pregnancy profoundly alters maternal physiology. The heart hypertrophies during pregnancy, but its metabolic adaptations, are not well understood.
Objective: To determine the mechanisms underlying cardiac substrate use during pregnancy.
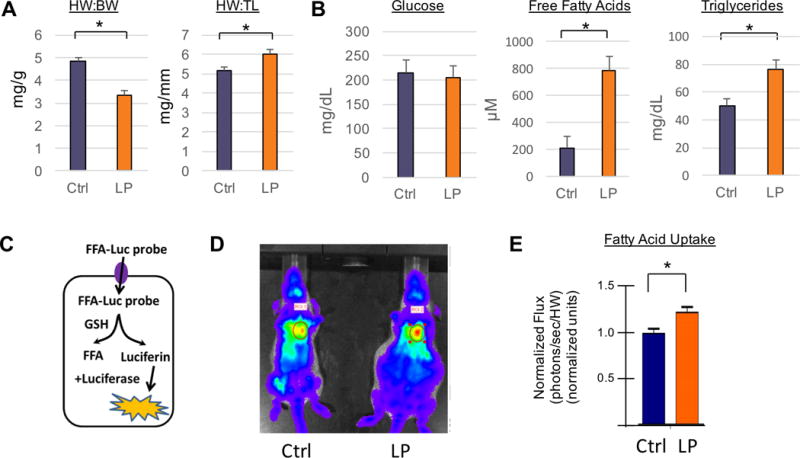
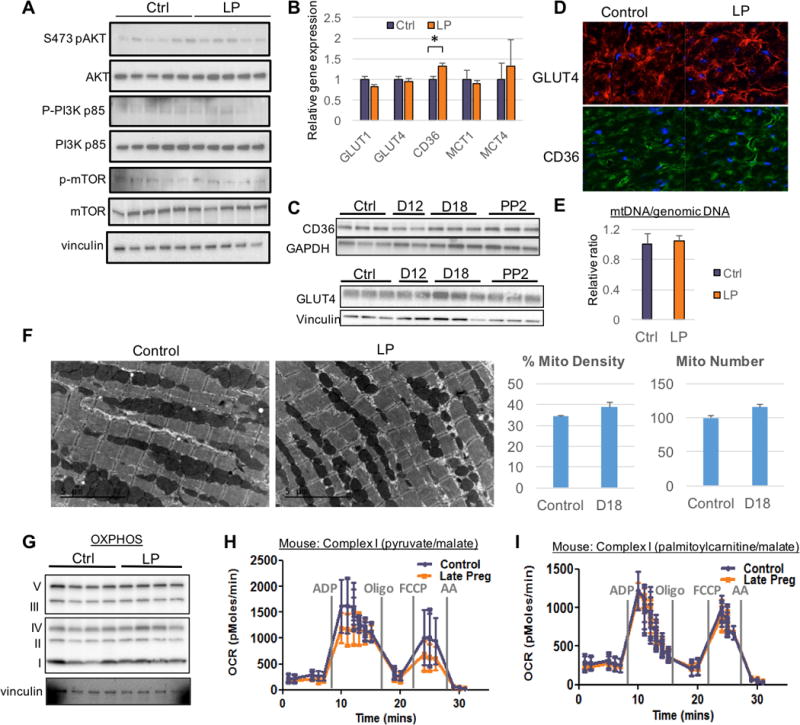
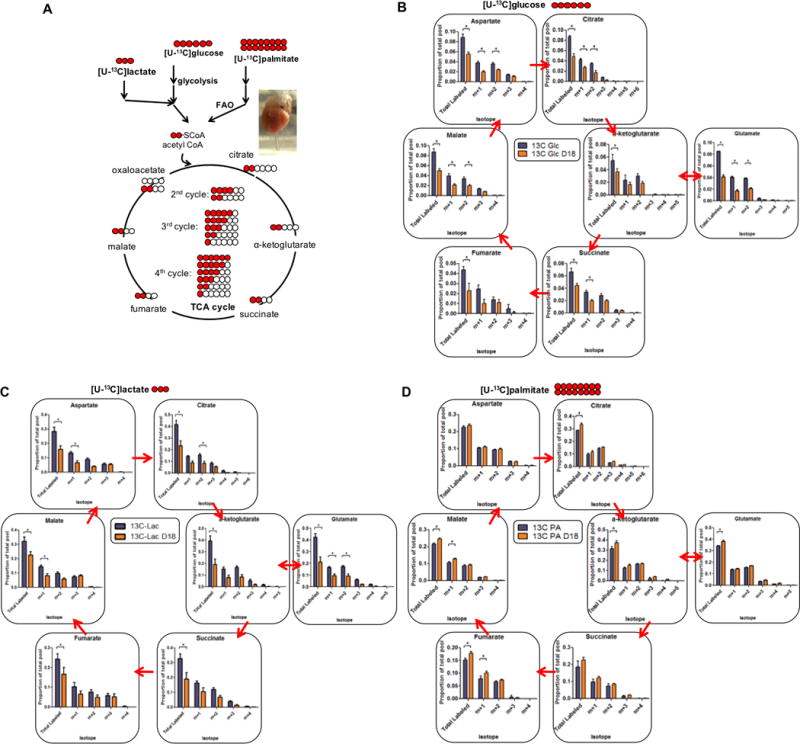
Methods and results: We use here 13C glucose, 13C lactate, and 13C fatty acid tracing analyses to show that hearts in late pregnant mice increase fatty acid uptake and oxidation into the tricarboxylic acid cycle, while reducing glucose and lactate oxidation. Mitochondrial quantity, morphology, and function do not seem altered. Insulin signaling seems intact, and the abundance and localization of the major fatty acid and glucose transporters, CD36 (cluster of differentiation 36) and GLUT4 (glucose transporter type 4), are also unchanged. Rather, we find that the pregnancy hormone progesterone induces PDK4 (pyruvate dehydrogenase kinase 4) in cardiomyocytes and that elevated PDK4 levels in late pregnancy lead to inhibition of PDH (pyruvate dehydrogenase) and pyruvate flux into the tricarboxylic acid cycle. Blocking PDK4 reverses the metabolic changes seen in hearts in late pregnancy.
Conclusions: Taken together, these data indicate that the hormonal environment of late pregnancy promotes metabolic remodeling in the heart at the level of PDH, rather than at the level of insulin signaling.
Keywords: glucose; heart; metabolism; mitochondria; pregnancy; progesterone.
© 2017 American Heart Association, Inc.
Figures

Comment in
-
Decreased Maternal Cardiac Glucose Oxidation: Taking One for the Fetus.Circ Res. 2017 Dec 8;121(12):1299-1301. doi: 10.1161/CIRCRESAHA.117.312098. Circ Res. 2017. PMID: 29217705 No abstract available.
References
-
- Lain KY, Catalano PM. Metabolic changes in pregnancy. Clin Obstet Gynecol. 2007;50:938–48. - PubMed
-
- Barbour LA, McCurdy CE, Hernandez TL, Kirwan JP, Catalano PM, Friedman JE. Cellular mechanisms for insulin resistance in normal pregnancy and gestational diabetes. Diabetes care. 2007;30(Suppl 2):S112–9. - PubMed
-
- Kirwan JP, Hauguel-De Mouzon S, Lepercq J, Challier JC, Huston-Presley L, Friedman JE, Kalhan SC, Catalano PM. TNF-alpha is a predictor of insulin resistance in human pregnancy. Diabetes. 2002;51:2207–13. - PubMed
-
- Melchiorre K, Sharma R, Thilaganathan B. Cardiac structure and function in normal pregnancy. Curr Opin Obstet Gynecol. 2012;24:413–21. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
