

Coordinated increase of nuclear tension and lamin-A with matrix stiffness outcompetes lamin-B receptor that favors soft tissue phenotypes
- PMID: 28931598
- PMCID: PMC5687034
- DOI: 10.1091/mbc.E17-06-0393
Coordinated increase of nuclear tension and lamin-A with matrix stiffness outcompetes lamin-B receptor that favors soft tissue phenotypes
Abstract
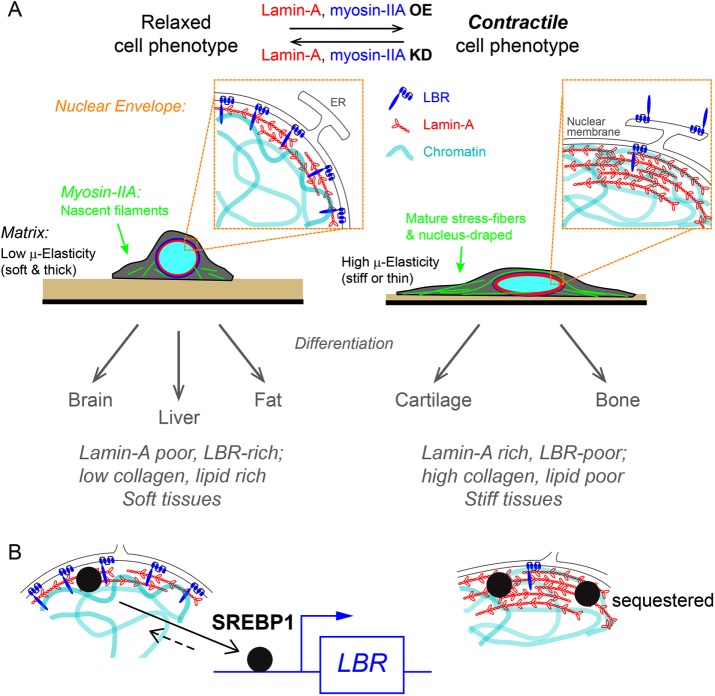
Matrix stiffness that is sensed by a cell or measured by a purely physical probe reflects the intrinsic elasticity of the matrix and also how thick or thin the matrix is. Here, mesenchymal stem cells (MSCs) and their nuclei spread in response to thickness-corrected matrix microelasticity, with increases in nuclear tension and nuclear stiffness resulting from increases in myosin-II and lamin-A,C. Linearity between the widely varying projected area of a cell and its nucleus across many matrices, timescales, and myosin-II activity levels indicates a constant ratio of nucleus-to-cell volume, despite MSCs' lineage plasticity. Nuclear envelope fluctuations are suppressed on the stiffest matrices, and fluctuation spectra reveal a high nuclear tension that matches trends from traction force microscopy and from increased lamin-A,C. Transcriptomes of many diverse tissues and MSCs further show that lamin-A,C's increase with tissue or matrix stiffness anti-correlates with lamin-B receptor (LBR), which contributes to lipid/sterol biosynthesis. Adipogenesis (a soft lineage) indeed increases LBR:lamin-A,C protein stoichiometry in MSCs versus osteogenesis (stiff). The two factors compete for lamin-B in response to matrix elasticity, knockdown, myosin-II inhibition, and even constricted migration that disrupts and segregates lamins in situ. Matrix stiffness-driven contractility thus tenses the nucleus to favor lamin-A,C accumulation and suppress soft tissue phenotypes.
© 2017 Buxboim et al. This article is distributed by The American Society for Cell Biology under license from the author(s). Two months after publication it is available to the public under an Attribution–Noncommercial–Share Alike 3.0 Unported Creative Commons License (http://creativecommons.org/licenses/by-nc-sa/3.0).
Figures
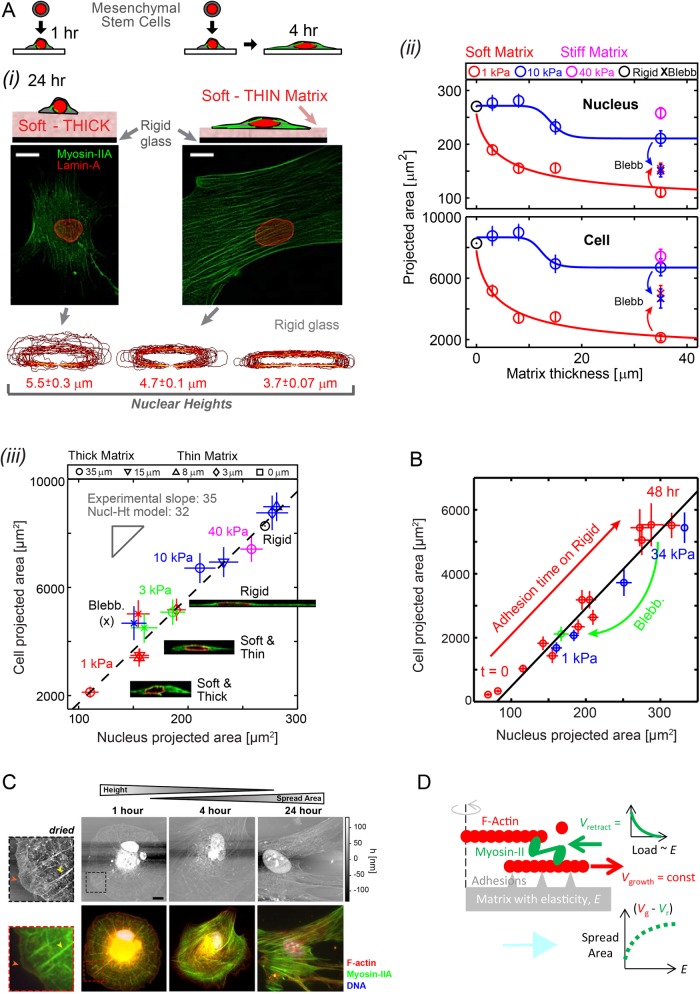
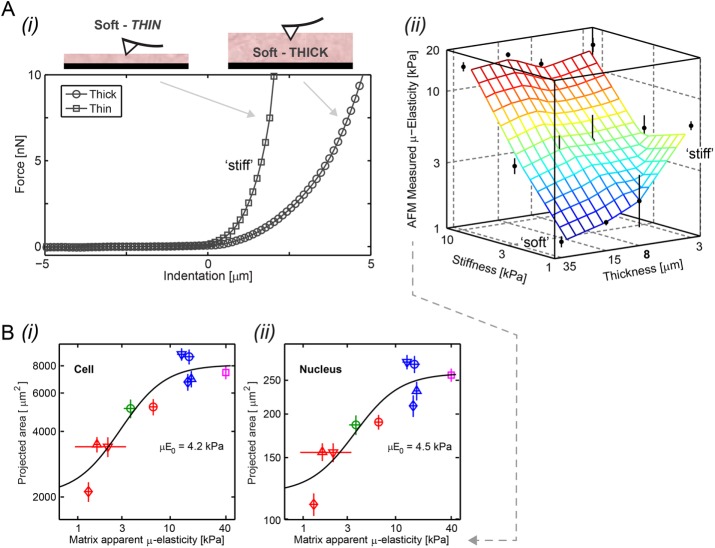
 , with an exponent of cooperativity n = 0.5. The transition between soft and stiff matrices is set by Eµ, amounting to 4–4.5 kPa, thus discriminating between compliant tissues such as brain, marrow, and fat and stiffer tissues such as muscle, cartilage, and bone.
, with an exponent of cooperativity n = 0.5. The transition between soft and stiff matrices is set by Eµ, amounting to 4–4.5 kPa, thus discriminating between compliant tissues such as brain, marrow, and fat and stiffer tissues such as muscle, cartilage, and bone.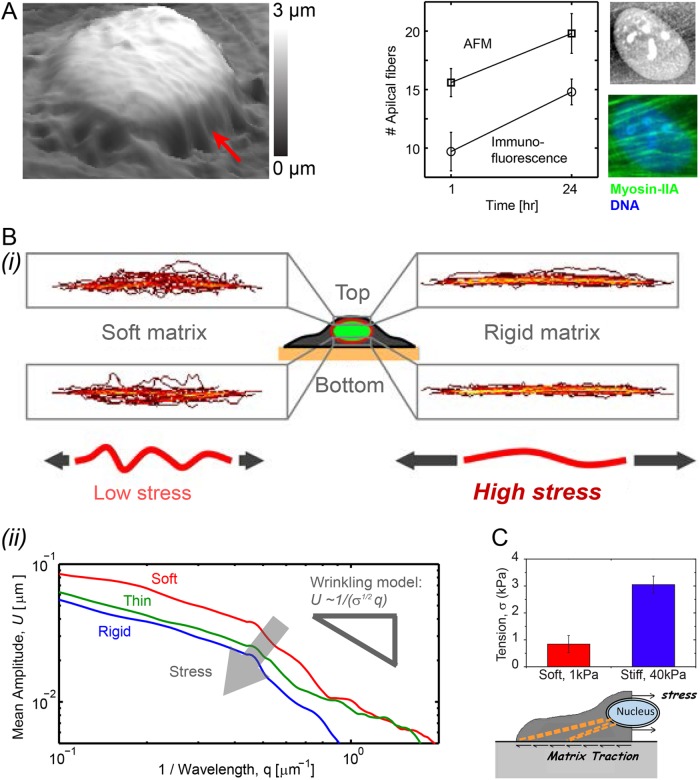
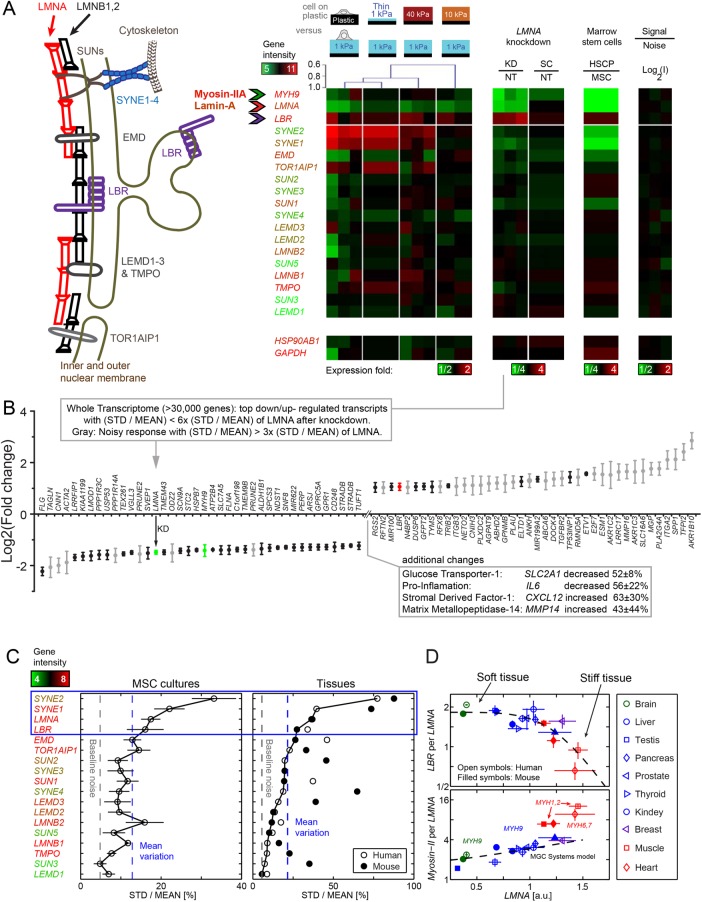
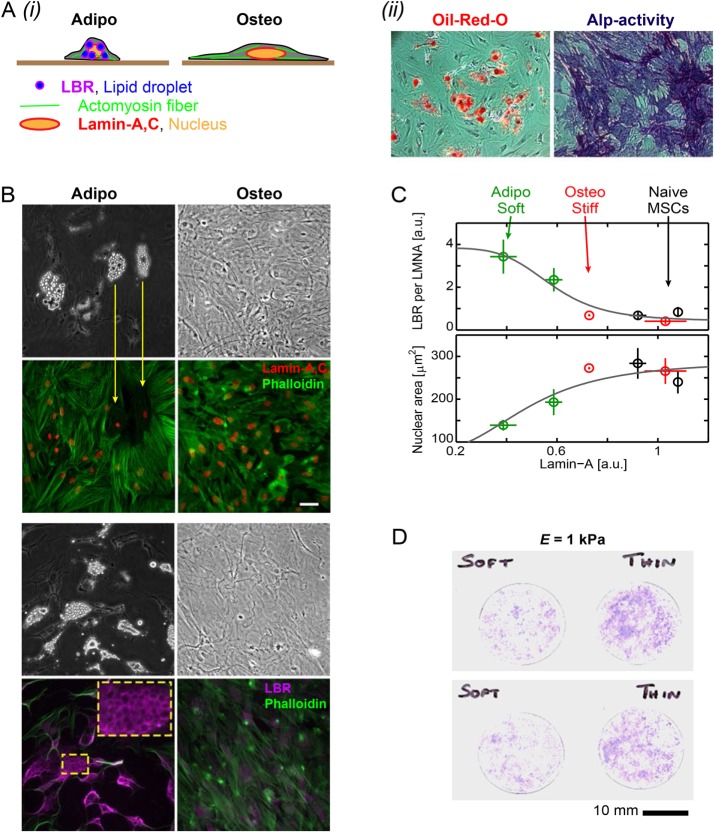
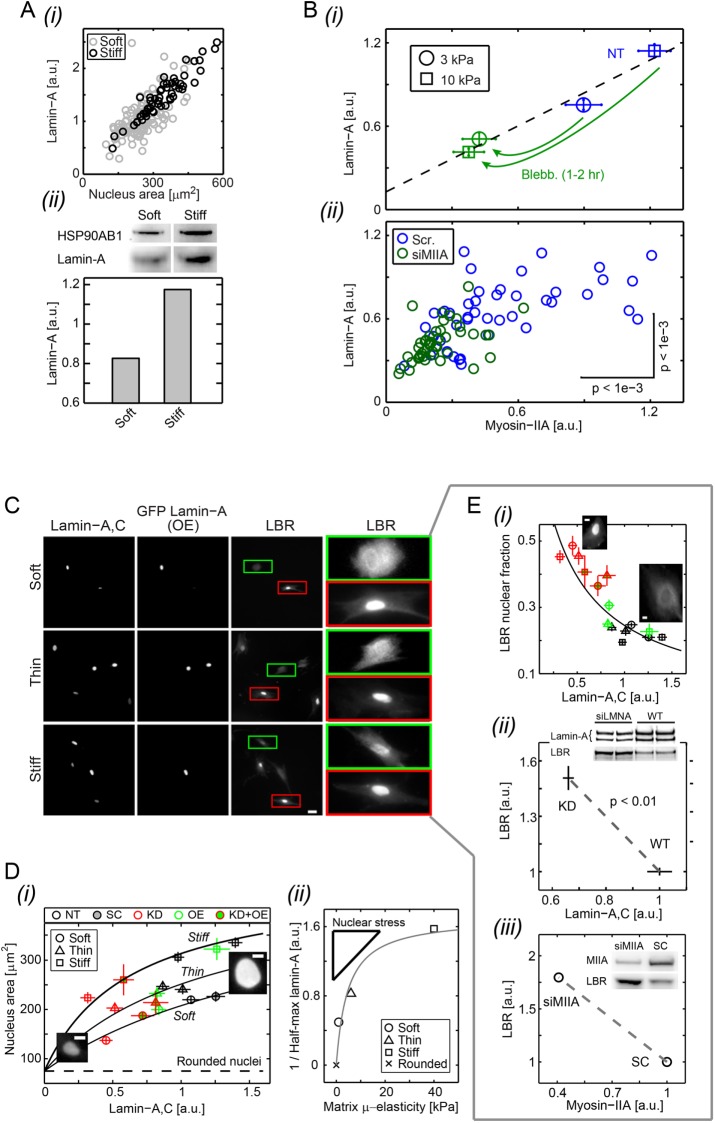
 intersect at x = 0, which corresponds to the projected area of rounded nuclei in cells in suspension. (ii) As A0 and β are the same across matrix fits and K is the lamin-A,C level for half-maximum nuclear stretching, 1/K relates to nuclear stress σ and increases with matrix μ-elasticity. (Ei) LBR nuclear fraction is anti-correlated with lamin-A,C across the various matrix conditions (legend in panel Di), and fits a competitive binding model. Overall protein levels of LBR are enhanced by knockdown of either lamin-A,C (i, siLMNA) or myosin-IIA (iii, siMIIA). Cell extract volumes were calibrated using Bradford total protein assay (i) or based on the immunoblotting intensities of HSP90AB1 housekeeping protein (ii). Average ± SEM, n > 25 cells. Scale bars = 10 µm.
intersect at x = 0, which corresponds to the projected area of rounded nuclei in cells in suspension. (ii) As A0 and β are the same across matrix fits and K is the lamin-A,C level for half-maximum nuclear stretching, 1/K relates to nuclear stress σ and increases with matrix μ-elasticity. (Ei) LBR nuclear fraction is anti-correlated with lamin-A,C across the various matrix conditions (legend in panel Di), and fits a competitive binding model. Overall protein levels of LBR are enhanced by knockdown of either lamin-A,C (i, siLMNA) or myosin-IIA (iii, siMIIA). Cell extract volumes were calibrated using Bradford total protein assay (i) or based on the immunoblotting intensities of HSP90AB1 housekeeping protein (ii). Average ± SEM, n > 25 cells. Scale bars = 10 µm.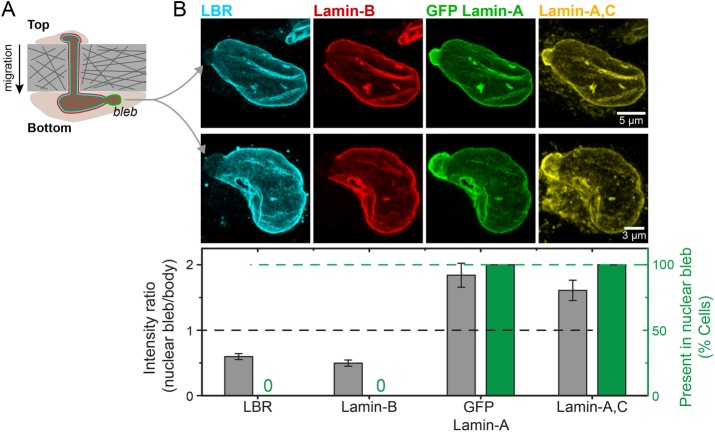
References
-
- Balcioglu HE, van Hoorn H, Donato DM, Schmidt T, Danen EH. The integrin expression profile modulates orientation and dynamics of force transmission at cell matrix adhesions. J Cell Sci. 2015;128:1316–1326. - PubMed
-
- Ben-Ze’ev A, Farmer SR, Penman S. Protein synthesis requires cell-surface contact while nuclear events respond to cell shape in anchorage-dependent fibroblasts. Cell. 1980;21:365–372. - PubMed
-
- Bjorkhem I, Meaney S. Brain cholesterol: long secret life behind a barrier. Arterioscler Thromb Vasc Biol. 2004;24:806–815. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
