

Connexins in Cardiovascular and Neurovascular Health and Disease: Pharmacological Implications
- PMID: 28931622
- PMCID: PMC5612248
- DOI: 10.1124/pr.115.012062
Connexins in Cardiovascular and Neurovascular Health and Disease: Pharmacological Implications
Abstract
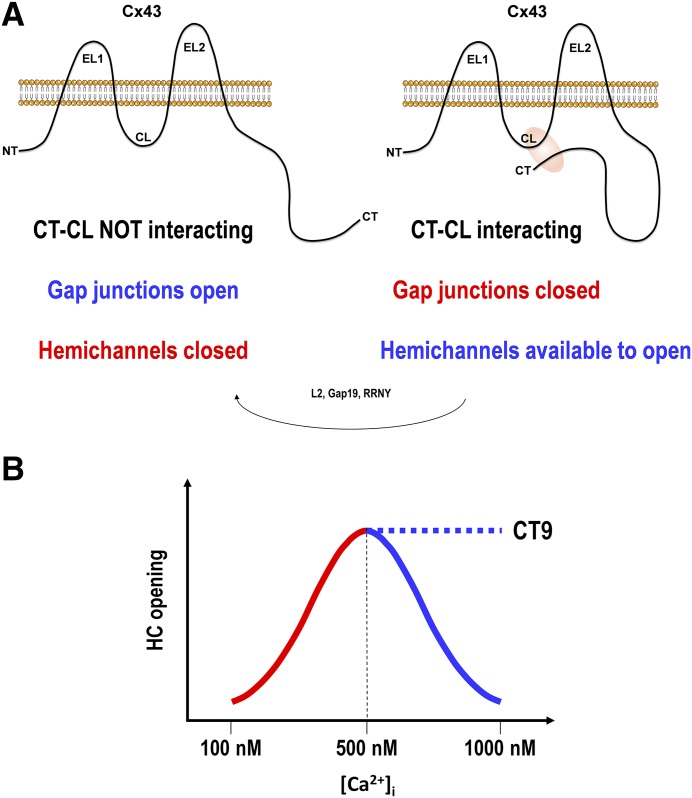
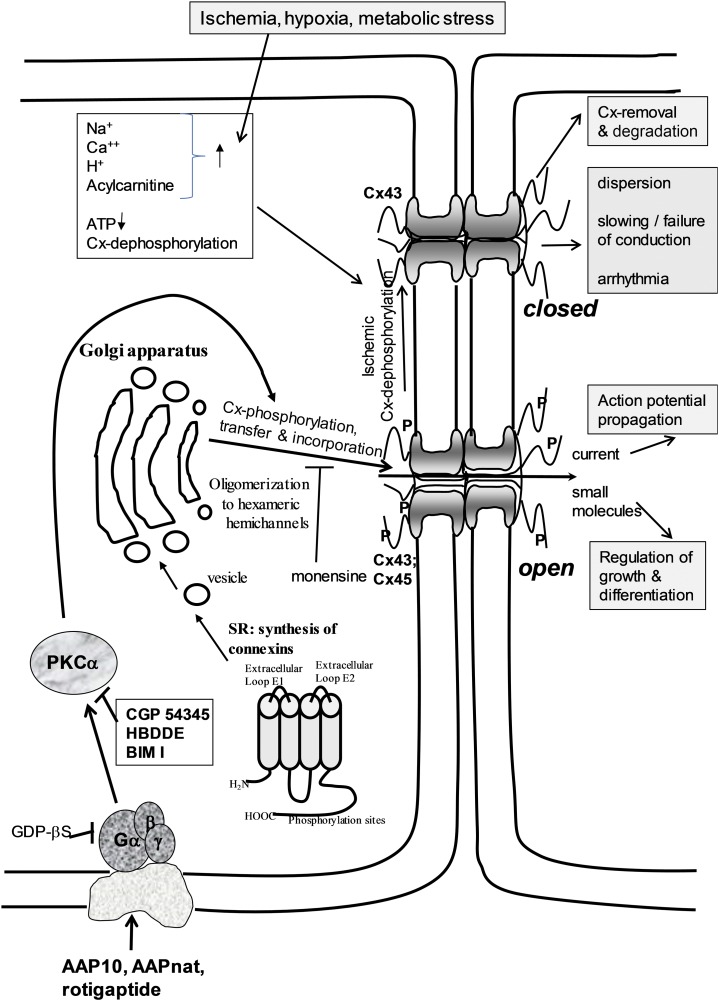
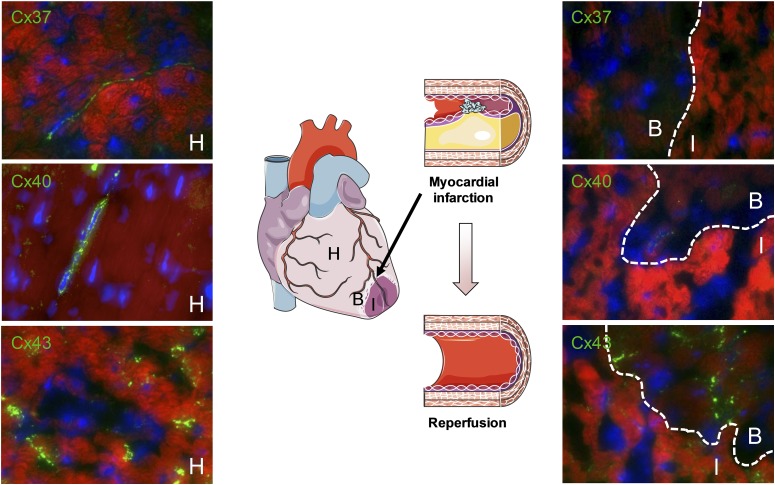
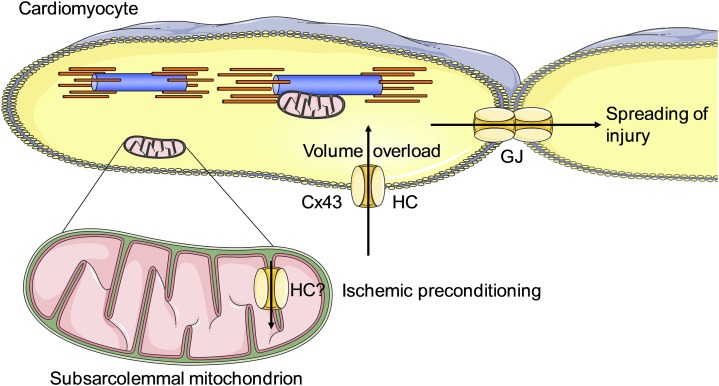
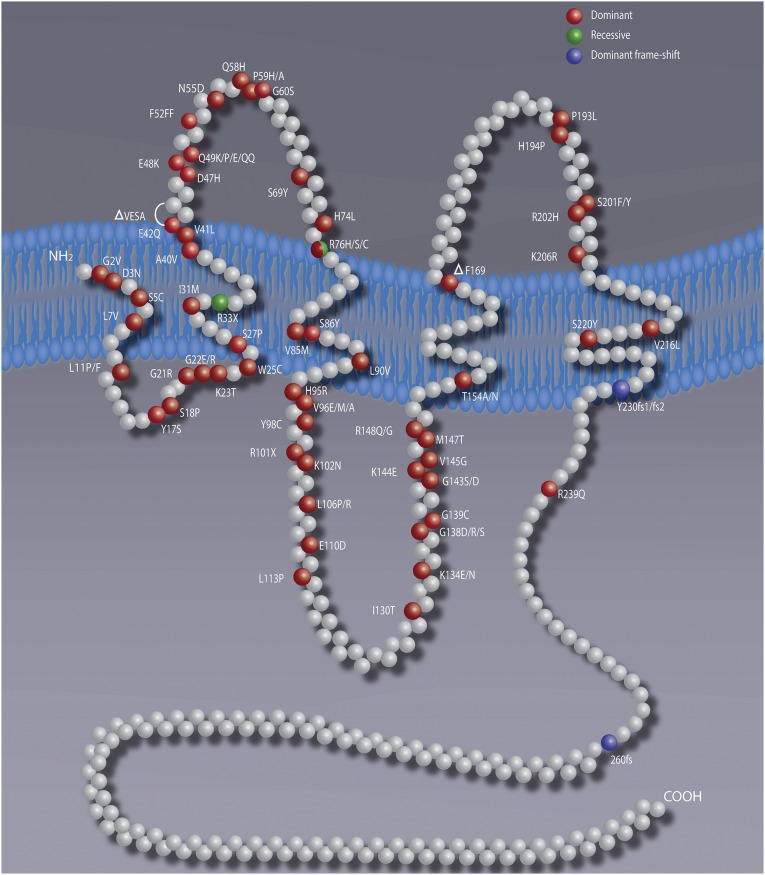
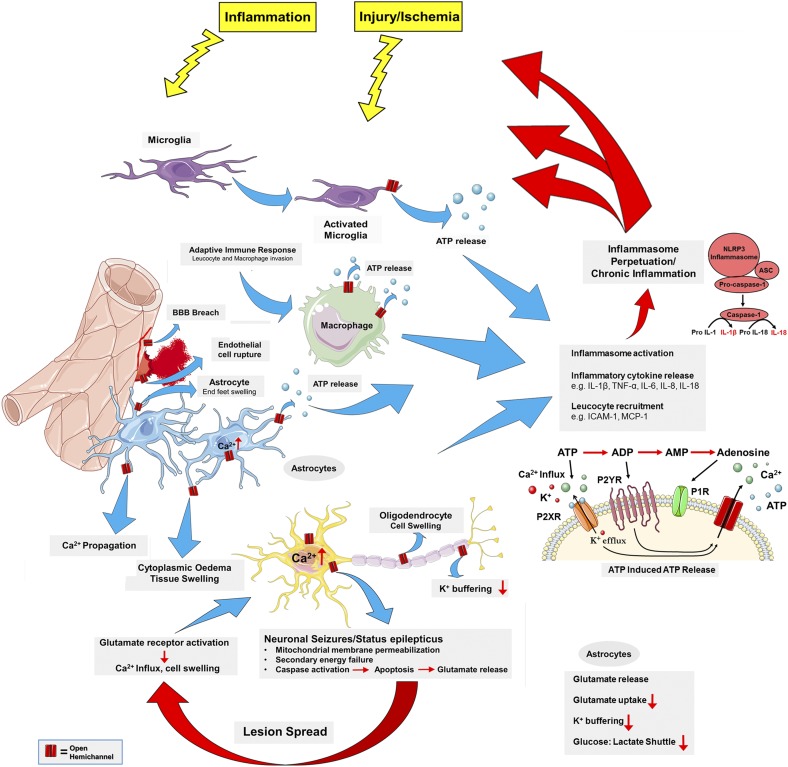
Connexins are ubiquitous channel forming proteins that assemble as plasma membrane hemichannels and as intercellular gap junction channels that directly connect cells. In the heart, gap junction channels electrically connect myocytes and specialized conductive tissues to coordinate the atrial and ventricular contraction/relaxation cycles and pump function. In blood vessels, these channels facilitate long-distance endothelial cell communication, synchronize smooth muscle cell contraction, and support endothelial-smooth muscle cell communication. In the central nervous system they form cellular syncytia and coordinate neural function. Gap junction channels are normally open and hemichannels are normally closed, but pathologic conditions may restrict gap junction communication and promote hemichannel opening, thereby disturbing a delicate cellular communication balance. Until recently, most connexin-targeting agents exhibited little specificity and several off-target effects. Recent work with peptide-based approaches has demonstrated improved specificity and opened avenues for a more rational approach toward independently modulating the function of gap junctions and hemichannels. We here review the role of connexins and their channels in cardiovascular and neurovascular health and disease, focusing on crucial regulatory aspects and identification of potential targets to modify their function. We conclude that peptide-based investigations have raised several new opportunities for interfering with connexins and their channels that may soon allow preservation of gap junction communication, inhibition of hemichannel opening, and mitigation of inflammatory signaling.
Copyright © 2017 by The American Society for Pharmacology and Experimental Therapeutics.
Figures


Similar articles
-
Emerging issues of connexin channels: biophysics fills the gap.Q Rev Biophys. 2001 Aug;34(3):325-472. doi: 10.1017/s0033583501003705. Q Rev Biophys. 2001. PMID: 11838236 Review.
-
Role of Non-Myocyte Gap Junctions and Connexin Hemichannels in Cardiovascular Health and Disease: Novel Therapeutic Targets?Int J Mol Sci. 2018 Mar 15;19(3):866. doi: 10.3390/ijms19030866. Int J Mol Sci. 2018. PMID: 29543751 Free PMC article. Review.
-
At the cross-point of connexins, calcium, and ATP: blocking hemichannels inhibits vasoconstriction of rat small mesenteric arteries.Cardiovasc Res. 2017 Feb;113(2):195-206. doi: 10.1093/cvr/cvw215. Epub 2016 Sep 27. Cardiovasc Res. 2017. PMID: 27677282
-
Connexin hemichannels as candidate targets for cardioprotective and anti-arrhythmic treatments.J Clin Invest. 2023 Mar 15;133(6):e168117. doi: 10.1172/JCI168117. J Clin Invest. 2023. PMID: 36919695 Free PMC article. Review.
-
Gap junction- and hemichannel-independent actions of connexins.Biochim Biophys Acta. 2005 Jun 10;1711(2):208-14. doi: 10.1016/j.bbamem.2004.10.001. Epub 2004 Oct 22. Biochim Biophys Acta. 2005. PMID: 15955305 Free PMC article. Review.
Cited by
-
Hypertensive Nephropathy: Unveiling the Possible Involvement of Hemichannels and Pannexons.Int J Mol Sci. 2022 Dec 14;23(24):15936. doi: 10.3390/ijms232415936. Int J Mol Sci. 2022. PMID: 36555574 Free PMC article. Review.
-
Cross-Activation of Hemichannels/Gap Junctions and Immunoglobulin-Like Domains in Innate-Adaptive Immune Responses.Front Immunol. 2022 Jul 15;13:882706. doi: 10.3389/fimmu.2022.882706. eCollection 2022. Front Immunol. 2022. PMID: 35911693 Free PMC article. Review.
-
Permeation of Molecules through Astroglial Connexin 43 Hemichannels Is Modulated by Cytokines with Parameters Depending on the Permeant Species.Int J Mol Sci. 2020 Jun 1;21(11):3970. doi: 10.3390/ijms21113970. Int J Mol Sci. 2020. PMID: 32492823 Free PMC article.
-
Connexins: Synthesis, Post-Translational Modifications, and Trafficking in Health and Disease.Int J Mol Sci. 2018 Apr 26;19(5):1296. doi: 10.3390/ijms19051296. Int J Mol Sci. 2018. PMID: 29701678 Free PMC article. Review.
-
Measurement of Ca2+ Uptake Through Connexin Hemichannels.Methods Mol Biol. 2024;2801:97-109. doi: 10.1007/978-1-0716-3842-2_8. Methods Mol Biol. 2024. PMID: 38578416
References
-
- Abascal F, Zardoya R. (2013) Evolutionary analyses of gap junction protein families. Biochim Biophys Acta 1828:4–14. - PubMed
-
- Abcouwer SF, Lin CM, Wolpert EB, Shanmugam S, Schaefer EW, Freeman WM, Barber AJ, Antonetti DA. (2010) Effects of ischemic preconditioning and bevacizumab on apoptosis and vascular permeability following retinal ischemia-reperfusion injury. Invest Ophthalmol Vis Sci 51:5920–5933. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
