

Zika Virus Encoding Nonglycosylated Envelope Protein Is Attenuated and Defective in Neuroinvasion
- PMID: 28931684
- PMCID: PMC5686755
- DOI: 10.1128/JVI.01348-17
Zika Virus Encoding Nonglycosylated Envelope Protein Is Attenuated and Defective in Neuroinvasion
Abstract
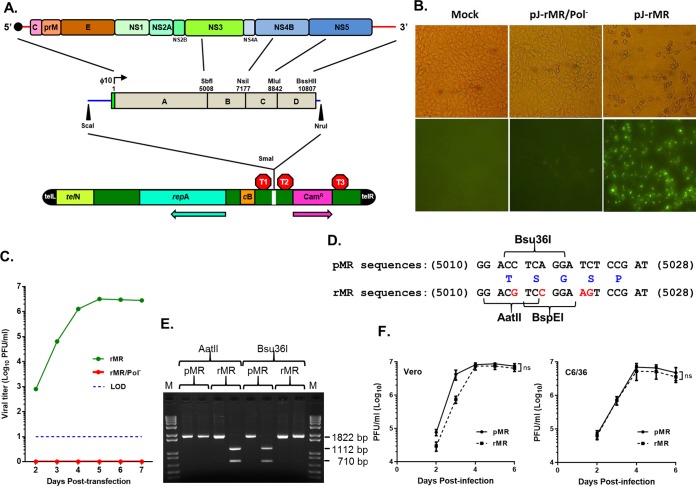
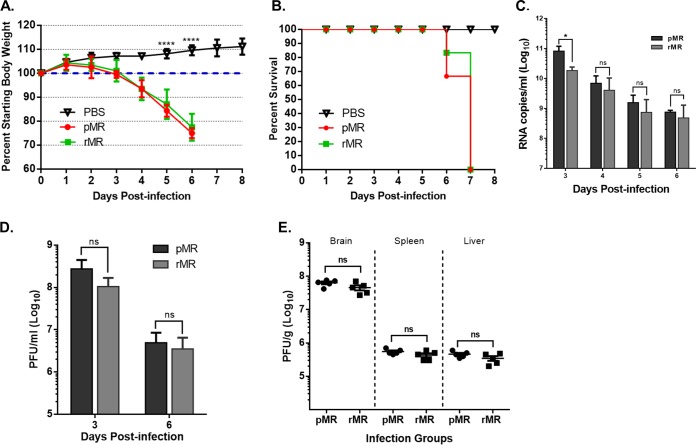
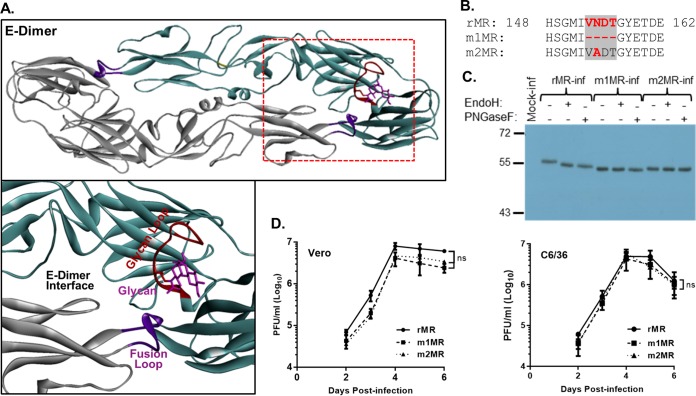
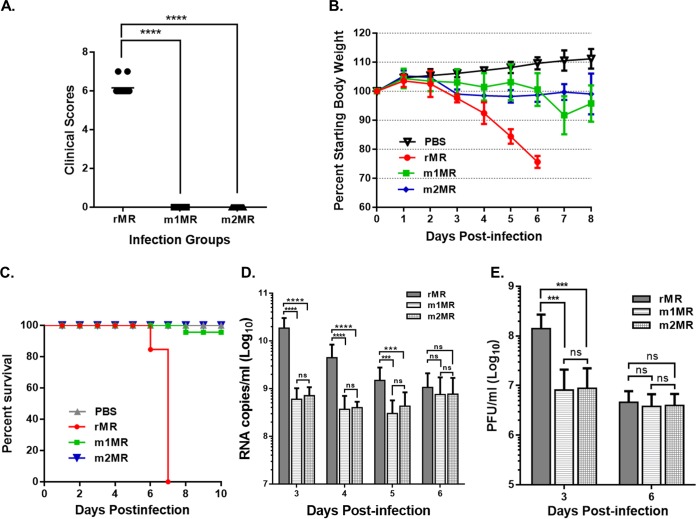
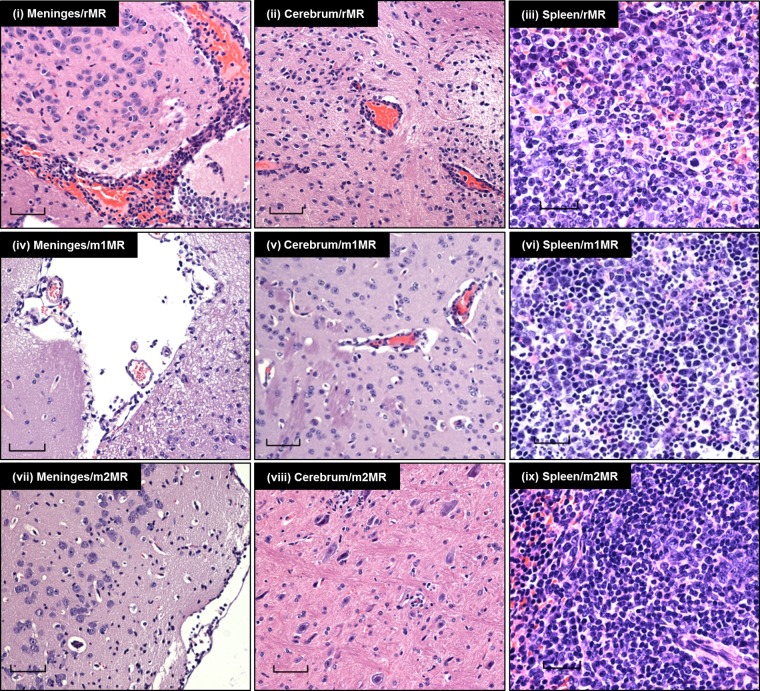
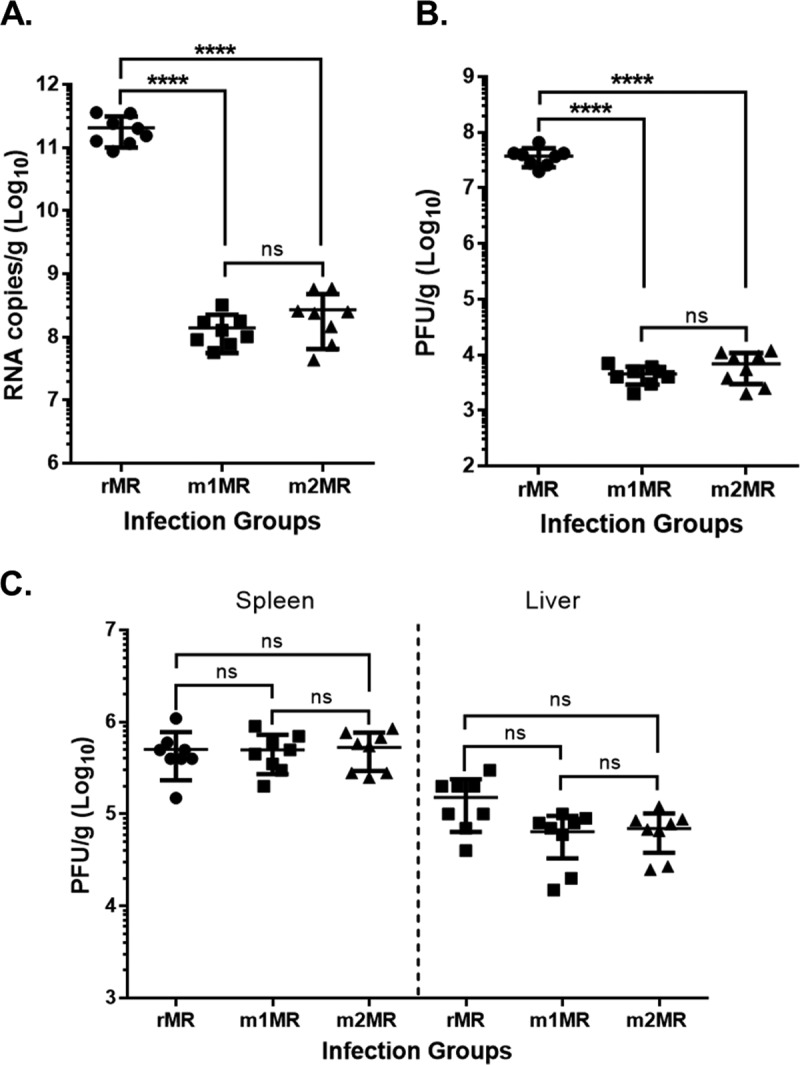
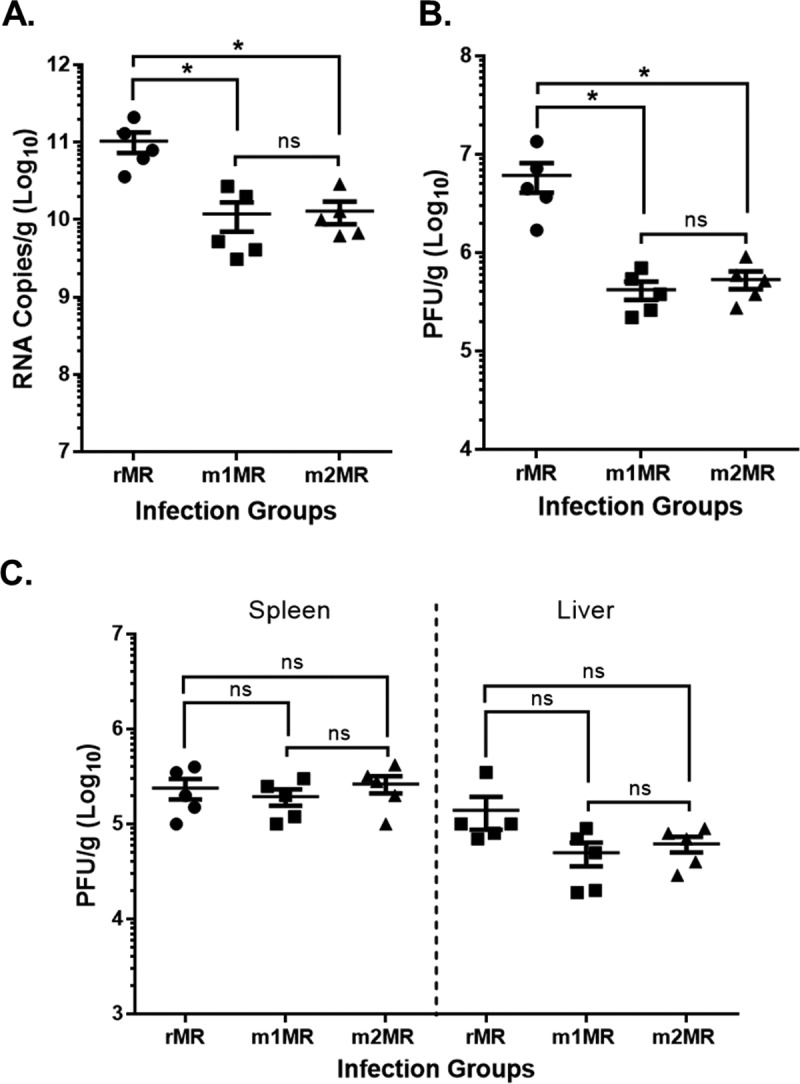
Zika virus (ZIKV), a mosquito-transmitted flavivirus responsible for sporadic outbreaks of mild and febrile illness in Africa and Asia, reemerged in the last decade causing serious human diseases, including microcephaly, congenital malformations, and Guillain-Barré syndrome. Although genomic and phylogenetic analyses suggest that genetic evolution may have led to the enhanced virulence of ZIKV, experimental evidence supporting the role of specific genetic changes in virulence is currently lacking. One sequence motif, VNDT, containing an N-linked glycosylation site in the envelope (E) protein, is polymorphic; it is absent in many of the African isolates but present in all isolates from the recent outbreaks. In the present study, we investigated the roles of this sequence motif and glycosylation of the E protein in the pathogenicity of ZIKV. We first constructed a stable full-length cDNA clone of ZIKV in a novel linear vector from which infectious virus was recovered. The recombinant ZIKV generated from the infectious clone, which contains the VNDT motif, is highly pathogenic and causes lethality in a mouse model. In contrast, recombinant viruses from which the VNDT motif is deleted or in which the N-linked glycosylation site is mutated by single-amino-acid substitution are highly attenuated and nonlethal. The mutant viruses replicate poorly in the brains of infected mice when inoculated subcutaneously but replicate well following intracranial inoculation. Our findings provide the first evidence that N-linked glycosylation of the E protein is an important determinant of ZIKV virulence and neuroinvasion.IMPORTANCE The recent emergence of Zika virus (ZIKV) in the Americas has caused major worldwide public health concern. The virus appears to have gained significant pathogenicity, causing serious human diseases, including microcephaly and Guillain-Barré syndrome. The factors responsible for the emergence of pathogenic ZIKV are not understood at this time, although genetic changes have been shown to facilitate virus transmission. All isolates from the recent outbreaks contain an N-linked glycosylation site within the viral envelope (E) protein, whereas many isolates of the African lineage virus lack this site. To elucidate the functional significance of glycosylation in ZIKV pathogenicity, recombinant ZIKVs from infectious clones with or without the glycan on the E protein were generated. ZIKVs lacking the glycan were highly attenuated for the ability to cause mortality in a mouse model and were severely compromised for neuroinvasion. Our studies suggest glycosylation of the E protein is an important factor contributing to ZIKV pathogenicity.
Keywords: E protein glycosylation; Zika virus; attenuation; neuroinvasion.
Copyright © 2017 American Society for Microbiology.
Figures
References
-
- Duffy MR, Chen TH, Hancock WT, Powers AM, Kool JL, Lanciotti RS, Pretrick M, Marfel M, Holzbauer S, Dubray C, Guillaumot L, Griggs A, Bel M, Lambert AJ, Laven J, Kosoy O, Panella A, Biggerstaff BJ, Fischer M, Hayes EB. 2009. Zika virus outbreak on Yap Island, Federated States of Micronesia. N Engl J Med 360:2536–2543. doi: 10.1056/NEJMoa0805715. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
