

Netrin-1 Derived from the Ventricular Zone, but not the Floor Plate, Directs Hindbrain Commissural Axons to the Ventral Midline
- PMID: 28931893
- PMCID: PMC5607380
- DOI: 10.1038/s41598-017-12269-8
Netrin-1 Derived from the Ventricular Zone, but not the Floor Plate, Directs Hindbrain Commissural Axons to the Ventral Midline
Abstract
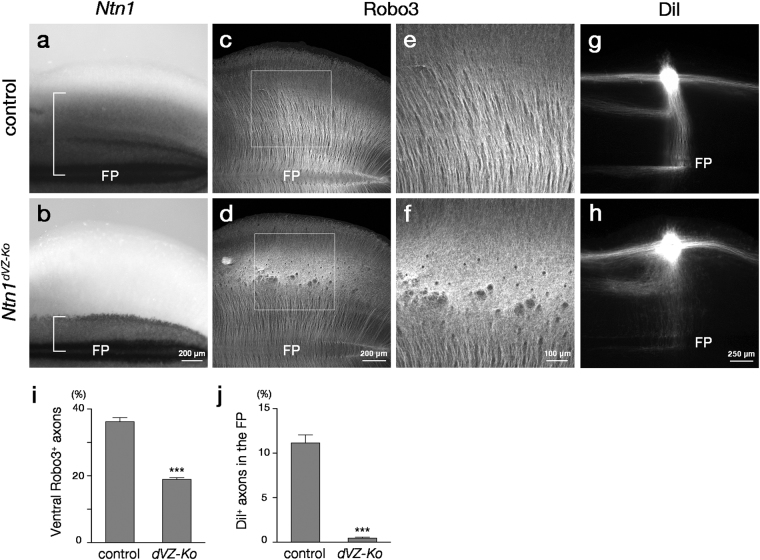
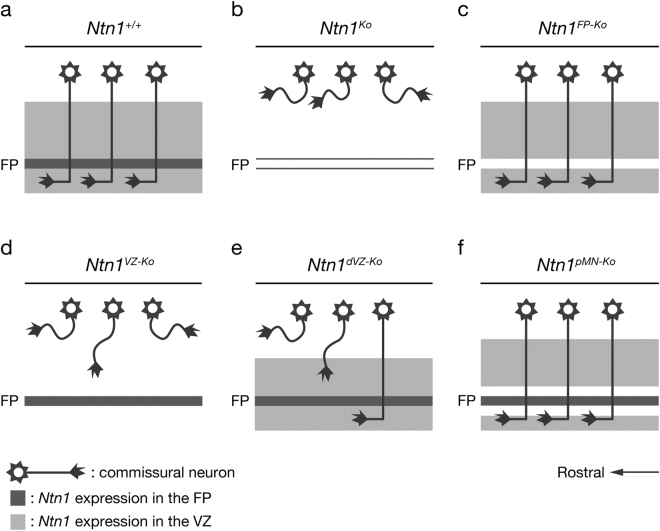
Netrin-1 (Ntn1) emanating from the ventral midline has been thought to act as a long-range diffusible chemoattractant for commissural axons (CAs). However, CAs still grow towards the midline in the absence of the floor plate (FP), a glial structure occupying the midline. Here, using genetically loss-of-function approaches in mice, we show that Ntn1 derived from the ventricular zone (VZ), but not the FP, is crucial for CA guidance in the mouse hindbrain. During the period of CA growth, Ntn1 is expressed in the ventral two-thirds of the VZ, in addition to the FP. Remarkably, deletion of Ntn1 from the VZ and even from the dorsal VZ highly disrupts CA guidance to the midline, whereas the deletion from the FP has little impact on it. We also show that the severities of CA guidance defects found in the Ntn1 conditional mutants were irrelevant to their FP long-range chemoattractive activities. Our results are incompatible with the prevailing view that Ntn1 is an FP-derived long-range diffusible chemoattractant for CAs, but suggest a novel mechanism that VZ-derived Ntn1 directs CAs to the ventral midline by its local actions.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures





Similar articles
-
Synergistic Activity of Floor-Plate- and Ventricular-Zone-Derived Netrin-1 in Spinal Cord Commissural Axon Guidance.Neuron. 2019 Feb 20;101(4):625-634.e3. doi: 10.1016/j.neuron.2018.12.024. Epub 2019 Jan 17. Neuron. 2019. PMID: 30661739
-
Long-Range Guidance of Spinal Commissural Axons by Netrin1 and Sonic Hedgehog from Midline Floor Plate Cells.Neuron. 2019 Feb 20;101(4):635-647.e4. doi: 10.1016/j.neuron.2018.12.025. Epub 2019 Jan 17. Neuron. 2019. PMID: 30661738
-
Floor-plate-derived netrin-1 is dispensable for commissural axon guidance.Nature. 2017 May 18;545(7654):350-354. doi: 10.1038/nature22331. Epub 2017 Apr 26. Nature. 2017. PMID: 28445456 Free PMC article.
-
The role of the floor plate in axon guidance.Annu Rev Neurosci. 1995;18:497-529. doi: 10.1146/annurev.ne.18.030195.002433. Annu Rev Neurosci. 1995. PMID: 7605072 Review.
-
A new model for netrin1 in commissural axon guidance.J Neurosci Res. 2018 Feb;96(2):247-252. doi: 10.1002/jnr.24117. Epub 2017 Jul 25. J Neurosci Res. 2018. PMID: 28742927 Review.
Cited by
-
A short isoform of the UNC-6/Netrin receptor UNC-5 is required for growth cone polarity and robust growth cone protrusion in Caenorhabditis elegans.bioRxiv [Preprint]. 2023 May 3:2023.05.02.539117. doi: 10.1101/2023.05.02.539117. bioRxiv. 2023. Update in: Front Cell Dev Biol. 2023 Aug 15;11:1240994. doi: 10.3389/fcell.2023.1240994. PMID: 37205526 Free PMC article. Updated. Preprint.
-
Rhes, a striatal enriched protein, regulates post-translational small-ubiquitin-like-modifier (SUMO) modification of nuclear proteins and alters gene expression.Cell Mol Life Sci. 2024 Apr 8;81(1):169. doi: 10.1007/s00018-024-05181-8. Cell Mol Life Sci. 2024. PMID: 38589732 Free PMC article.
-
TOM-1/tomosyn acts with the UNC-6/netrin receptor UNC-5 to inhibit growth cone protrusion in Caenorhabditis elegans.Development. 2023 Apr 1;150(7):dev201031. doi: 10.1242/dev.201031. Epub 2023 Apr 4. Development. 2023. PMID: 37014062 Free PMC article.
-
Short- and long-range roles of UNC-6/Netrin in dorsal-ventral axon guidance in vivo in Caenorhabditis elegans.bioRxiv [Preprint]. 2024 Apr 23:2024.04.23.590737. doi: 10.1101/2024.04.23.590737. bioRxiv. 2024. Update in: PLoS Genet. 2025 Jan 17;21(1):e1011526. doi: 10.1371/journal.pgen.1011526. PMID: 38712249 Free PMC article. Updated. Preprint.
-
Regulators of Rho GTPases in the Nervous System: Molecular Implication in Axon Guidance and Neurological Disorders.Int J Mol Sci. 2019 Mar 25;20(6):1497. doi: 10.3390/ijms20061497. Int J Mol Sci. 2019. PMID: 30934641 Free PMC article. Review.
References
-
- Cajal, S. R. Y. Histology of the nervous system of man and vertebrates. (Oxford University Press, USA, 1995).
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
