

Alpha TC1 and Beta-TC-6 genomic profiling uncovers both shared and distinct transcriptional regulatory features with their primary islet counterparts
- PMID: 28931935
- PMCID: PMC5607285
- DOI: 10.1038/s41598-017-12335-1
Alpha TC1 and Beta-TC-6 genomic profiling uncovers both shared and distinct transcriptional regulatory features with their primary islet counterparts
Abstract
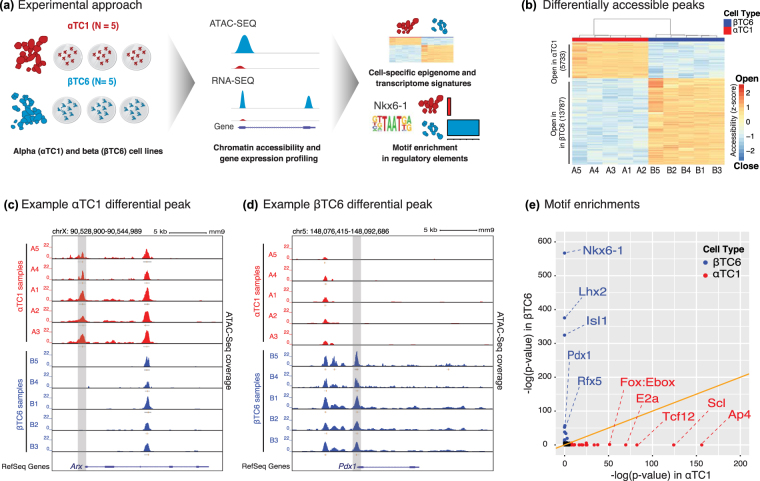
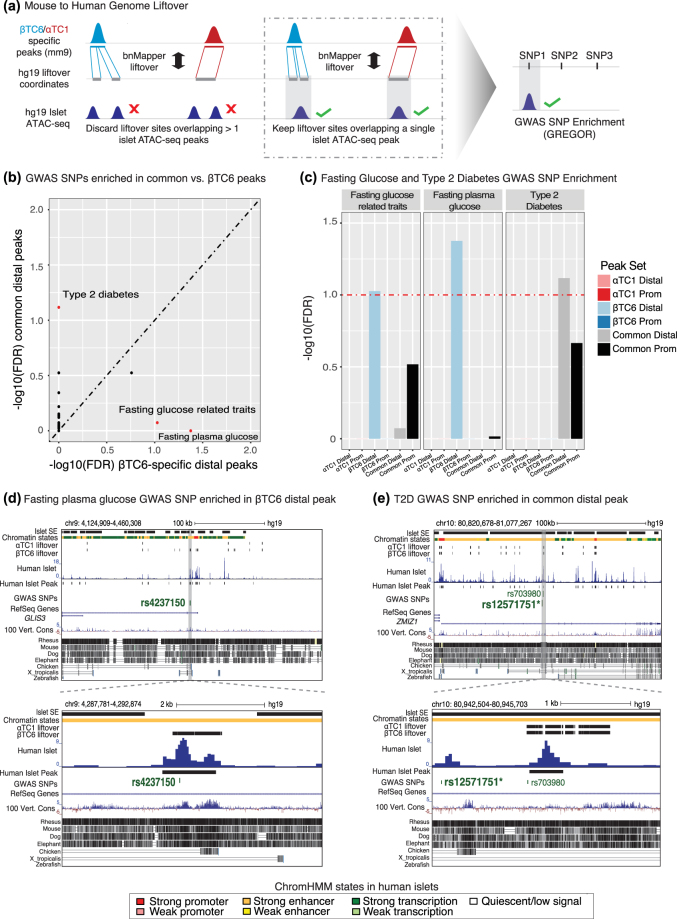
Alpha TC1 (αTC1) and Beta-TC-6 (βTC6) mouse islet cell lines are cellular models of islet (dys)function and type 2 diabetes (T2D). However, genomic characteristics of these cells, and their similarities to primary islet alpha and beta cells, are undefined. Here, we report the epigenomic (ATAC-seq) and transcriptomic (RNA-seq) landscapes of αTC1 and βTC6 cells. Each cell type exhibits hallmarks of its primary islet cell counterpart including cell-specific expression of beta (e.g., Pdx1) and alpha (e.g., Arx) cell transcription factors (TFs), and enrichment of binding motifs for these TFs in αTC1/βTC6 cis-regulatory elements. αTC1/βTC6 transcriptomes overlap significantly with the transcriptomes of primary mouse/human alpha and beta cells. Our data further indicate that ATAC-seq detects cell-specific regulatory elements for cell types comprising ≥ 20% of a mixed cell population. We identified αTC1/βTC6 cis-regulatory elements orthologous to those containing type 2 diabetes (T2D)-associated SNPs in human islets for 33 loci, suggesting these cells' utility to dissect T2D molecular genetics in these regions. Together, these maps provide important insights into the conserved regulatory architecture between αTC1/βTC6 and primary islet cells that can be leveraged in functional (epi)genomic approaches to dissect the genetic and molecular factors controlling islet cell identity and function.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures


References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
