

Neurofibromatosis type 1 alternative splicing is a key regulator of Ras/ERK signaling and learning behaviors in mice
- PMID: 28934393
- PMCID: PMC5886178
- DOI: 10.1093/hmg/ddx264
Neurofibromatosis type 1 alternative splicing is a key regulator of Ras/ERK signaling and learning behaviors in mice
Abstract
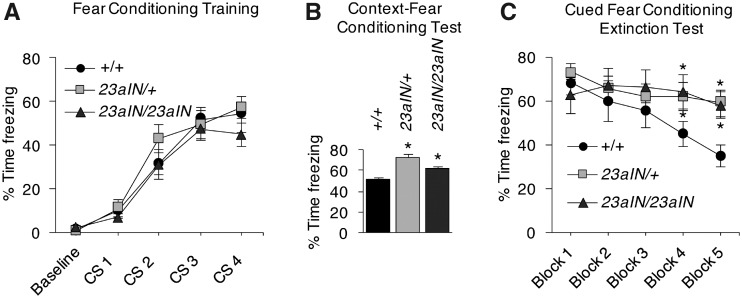
Appropriate activation of the Ras/extracellular signal-regulated kinase (ERK) protein signaling cascade within the brain is crucial for optimal learning and memory. One key regulator of this cascade is the Nf1 Ras GTPase activating protein (RasGAP), which attenuates Ras/ERK signaling by converting active Ras is bound to guanosine triphosphate, activating Ras into inactive Ras is bound to guanosine diphosphate, inactivating Ras. A previous study using embryonic stem cells and embryonic stem cell-derived neurons indicated that Nf1 RasGAP activity is modulated by the highly regulated alternative splicing of Nf1 exon 23a. In this study, we generated Nf123aIN/23aIN mice, in which the splicing signals surrounding Nf1 exon 23a were manipulated to increase exon inclusion. Nf123aIN/23aIN mice are viable and exon 23a inclusion approaches 100% in all tissues, including the brain, where the exon is normally almost completely skipped. Ras activation and phosphorylation of ERK1/2 downstream of Ras are both greatly increased in Nf123aIN/23aIN mouse brain lysates, confirming that exon 23a inclusion inhibits Nf1 RasGAP activity in vivo as it does in cultured cells. Consistent with the finding of altered Ras/ERK signaling in the brain, Nf123aIN/23aIN mice showed specific deficits in learning and memory compared with Nf1+/+ mice. Nf123aIN/23aIN mice performed poorly on the T-maze and Morris water maze tests, which measure short- and long-term spatial memory, respectively. In addition, Nf123aIN/23aIN mice showed abnormally elevated context-dependent fear and a diminished ability to extinguish a cued fear response, indicating defective associative fear learning. Therefore, the regulated alternative splicing of Nf1 is an important mechanism for fine-tuning Ras/ERK signaling as well as learning and memory in mice.
© The Author 2017. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures

Similar articles
-
Neurofibromatosis type 1 alternative splicing is a key regulator of Ras signaling in neurons.Mol Cell Biol. 2014 Jun;34(12):2188-97. doi: 10.1128/MCB.00019-14. Epub 2014 Apr 7. Mol Cell Biol. 2014. PMID: 24710274 Free PMC article.
-
Alternative splicing of the neurofibromatosis type I pre-mRNA.Biosci Rep. 2012 Apr 1;32(2):131-8. doi: 10.1042/BSR20110060. Biosci Rep. 2012. PMID: 22115364 Free PMC article. Review.
-
Alternative Splicing of Exon 23a in Neurofibromatosis Type 1 Pre-mRNA: Its Contribution to the Protein Structure and Function of Neurofibromin.Wiley Interdiscip Rev RNA. 2025 Jul-Aug;16(4):e70021. doi: 10.1002/wrna.70021. Wiley Interdiscip Rev RNA. 2025. PMID: 40812791 Free PMC article. Review.
-
Using antisense oligonucleotides for the physiological modulation of the alternative splicing of NF1 exon 23a during PC12 neuronal differentiation.Sci Rep. 2021 Feb 11;11(1):3661. doi: 10.1038/s41598-021-83152-w. Sci Rep. 2021. PMID: 33574490 Free PMC article.
-
Elucidating the impact of neurofibromatosis-1 germline mutations on neurofibromin function and dopamine-based learning.Hum Mol Genet. 2015 Jun 15;24(12):3518-28. doi: 10.1093/hmg/ddv103. Epub 2015 Mar 18. Hum Mol Genet. 2015. PMID: 25788518 Free PMC article.
Cited by
-
Quantitative Analysis of Alternative Pre-mRNA Splicing in Mouse Brain Sections Using RNA In Situ Hybridization Assay.J Vis Exp. 2018 Aug 26;(138):57889. doi: 10.3791/57889. J Vis Exp. 2018. PMID: 30199013 Free PMC article.
-
Allosteric modulation of NF1 GAP: Differential distributions of catalytically competent populations in loss-of-function and gain-of-function mutants.Protein Sci. 2025 Feb;34(2):e70042. doi: 10.1002/pro.70042. Protein Sci. 2025. PMID: 39840811 Free PMC article.
-
Mechanistic insights from animal models of neurofibromatosis type 1 cognitive impairment.Dis Model Mech. 2022 Aug 1;15(8):dmm049422. doi: 10.1242/dmm.049422. Epub 2022 Aug 29. Dis Model Mech. 2022. PMID: 36037004 Free PMC article. Review.
-
Assessment of Mosaicism and Detection of Cryptic Alleles in CRISPR/Cas9-Engineered Neurofibromatosis Type 1 and TP53 Mutant Porcine Models Reveals Overlooked Challenges in Precision Modeling of Human Diseases.Front Genet. 2021 Sep 23;12:721045. doi: 10.3389/fgene.2021.721045. eCollection 2021. Front Genet. 2021. PMID: 34630515 Free PMC article.
-
Neurobehavioral sex-related differences in Nf1+/- mice: female show a "camouflaging"-type behavior.Biol Sex Differ. 2023 Apr 26;14(1):24. doi: 10.1186/s13293-023-00509-8. Biol Sex Differ. 2023. PMID: 37101298 Free PMC article.
References
-
- Brambilla R., Gnesutta N., Minichiello L., White G., Roylance A.J., Herron C.E., Ramsey M., Wolfer D.P., Cestari V., Rossi-Arnaud C.. et al. (1997) A role for the Ras signalling pathway in synaptic transmission and long-term memory. Nature, 390, 281–286. - PubMed
-
- Komiyama N.H., Watabe A.M., Carlisle H.J., Porter K., Charlesworth P., Monti J., Strathdee D.J., O'Carroll C.M., Martin S.J., Morris R.G.. et al. (2002) SynGAP regulates ERK/MAPK signaling, synaptic plasticity, and learning in the complex with postsynaptic density 95 and NMDA receptor. J. Neurosci., 22, 9721–9732. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
