

Epstein-Barr Virus Type 2 Infects T Cells in Healthy Kenyan Children
- PMID: 28934430
- PMCID: PMC5853903
- DOI: 10.1093/infdis/jix363
Epstein-Barr Virus Type 2 Infects T Cells in Healthy Kenyan Children
Abstract
Background: The 2 strains of Epstein-Barr virus (EBV), EBV type 1 (EBV-1) and EBV-2, differ in latency genes, suggesting that they use distinct mechanisms to establish latency. We previously reported that EBV-2 infects T cells in vitro. In this study, we tested the possibility that EBV-2 infects T cells in vivo.
Methods: Purified T-cell fractions isolated from children positive for EBV-1 or EBV-2 and their mothers were examined for the presence of EBV and for EBV type.
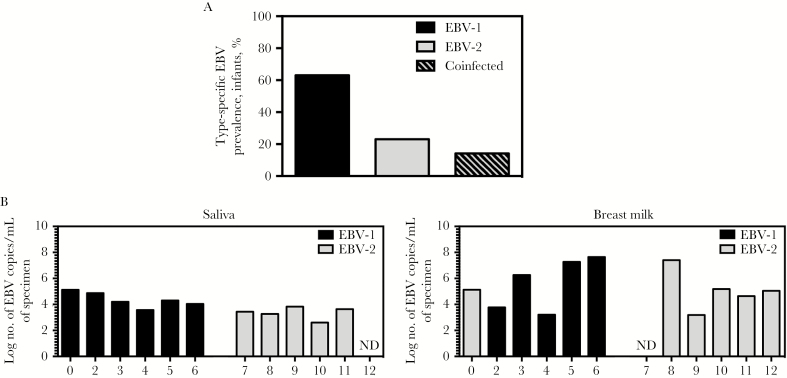
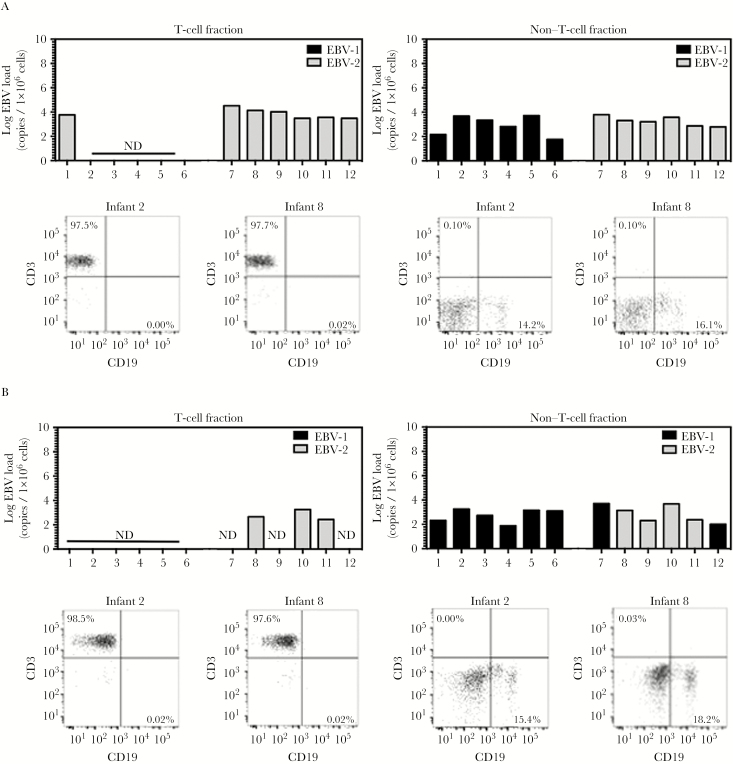
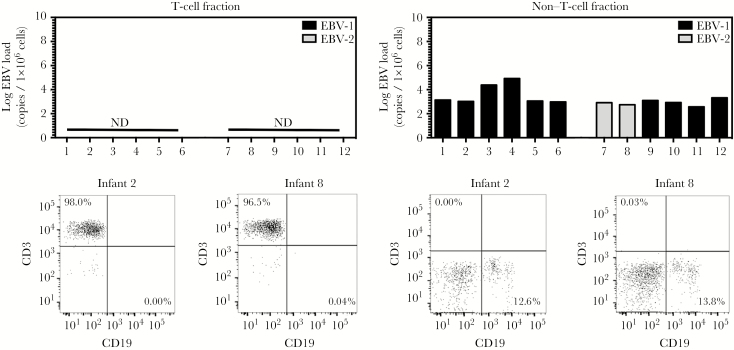
Results: We detected EBV-2 in all T-cell samples obtained from EBV-2-infected children at 12 months of age, with some children retaining EBV-2-positive T cells through 24 months of age, suggesting that EBV-2 persists in T cells. We were unable to detect EBV-2 in T-cell samples from mothers but could detect EBV-2 in samples of their breast milk and saliva.
Conclusions: These data suggest that EBV-2 uses T cells as an additional latency reservoir but that, over time, the frequency of infected T cells may drop below detectable levels. Alternatively, EBV-2 may establish a prolonged transient infection in the T-cell compartment. Collectively, these novel findings demonstrate that EBV-2 infects T cells in vivo and suggest EBV-2 may use the T-cell compartment to establish latency.
Keywords: Burkitt lymphoma; Epstein-Barr virus; T lymphocytes; cellular tropism.
© The Author 2017. Published by Oxford University Press for the Infectious Diseases Society of America. All rights reserved. For permissions, e-mail: journals.permissions@oup.com.
Figures
Similar articles
-
Epstein-Barr Virus Type 2 Infects T Cells and Induces B Cell Lymphomagenesis in Humanized Mice.J Virol. 2018 Oct 12;92(21):e00813-18. doi: 10.1128/JVI.00813-18. Print 2018 Nov 1. J Virol. 2018. PMID: 30089703 Free PMC article.
-
Breast Milk as a Potential Source of Epstein-Barr Virus Transmission Among Infants Living in a Malaria-Endemic Region of Kenya.J Infect Dis. 2015 Dec 1;212(11):1735-42. doi: 10.1093/infdis/jiv290. Epub 2015 May 17. J Infect Dis. 2015. PMID: 25985902 Free PMC article.
-
Epstein-Barr virus type 2 latently infects T cells, inducing an atypical activation characterized by expression of lymphotactic cytokines.J Virol. 2015 Feb;89(4):2301-12. doi: 10.1128/JVI.03001-14. Epub 2014 Dec 10. J Virol. 2015. PMID: 25505080 Free PMC article.
-
Regulation and dysregulation of Epstein-Barr virus latency: implications for the development of autoimmune diseases.Autoimmunity. 2008 May;41(4):298-328. doi: 10.1080/08916930802024772. Autoimmunity. 2008. PMID: 18432410 Review.
-
[Epstein-Barr virus penetration mechanism to T lymphoblastoid cell lines].Nihon Rinsho. 1997 Feb;55(2):404-8. Nihon Rinsho. 1997. PMID: 9046832 Review. Japanese.
Cited by
-
Epstein-Barr Virus (EBV) Genotypes Associated with the Immunopathological Profile of People Living with HIV-1: Immunological Aspects of Primary EBV Infection.Viruses. 2022 Jan 18;14(2):168. doi: 10.3390/v14020168. Viruses. 2022. PMID: 35215762 Free PMC article.
-
Epstein-Barr Virus Type 2 Infects T Cells and Induces B Cell Lymphomagenesis in Humanized Mice.J Virol. 2018 Oct 12;92(21):e00813-18. doi: 10.1128/JVI.00813-18. Print 2018 Nov 1. J Virol. 2018. PMID: 30089703 Free PMC article.
-
Peripheral T cell lymphomas: from the bench to the clinic.Nat Rev Cancer. 2020 Jun;20(6):323-342. doi: 10.1038/s41568-020-0247-0. Epub 2020 Apr 6. Nat Rev Cancer. 2020. PMID: 32249838 Review.
-
Epstein-Barr virus NK and T cell lymphoproliferative disease: report of a 2018 international meeting.Leuk Lymphoma. 2020 Apr;61(4):808-819. doi: 10.1080/10428194.2019.1699080. Epub 2019 Dec 13. Leuk Lymphoma. 2020. PMID: 31833428 Free PMC article. Review.
-
Epstein-Barr Virus Epidemiology, Serology, and Genetic Variability of LMP-1 Oncogene Among Healthy Population: An Update.Front Oncol. 2018 Jun 13;8:211. doi: 10.3389/fonc.2018.00211. eCollection 2018. Front Oncol. 2018. PMID: 29951372 Free PMC article. Review.
References
-
- Young LS, Yao QY, Rooney CM et al. . New type B isolates of Epstein-Barr virus from Burkitt’s lymphoma and from normal individuals in endemic areas. J Gen Virol 1987; 68 (Pt 11):2853–62. - PubMed
-
- Babcock GJ, Hochberg D, Thorley-Lawson AD. The expression pattern of Epstein-Barr virus latent genes in vivo is dependent upon the differentiation stage of the infected B cell. Immunity 2000; 13:497–506. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
