

Cohesin acetyltransferase Esco2 regulates SAC and kinetochore functions via maintaining H4K16 acetylation during mouse oocyte meiosis
- PMID: 28934466
- PMCID: PMC5766191
- DOI: 10.1093/nar/gkx563
Cohesin acetyltransferase Esco2 regulates SAC and kinetochore functions via maintaining H4K16 acetylation during mouse oocyte meiosis
Abstract
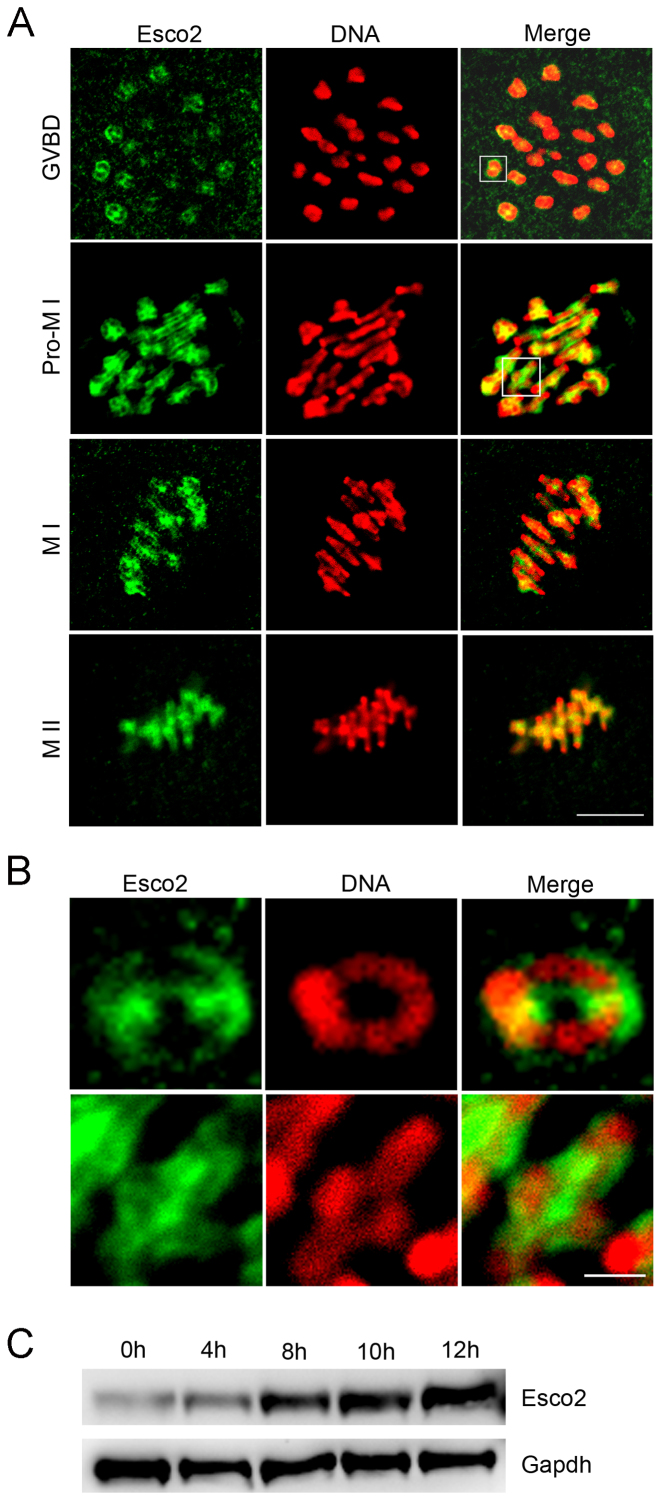
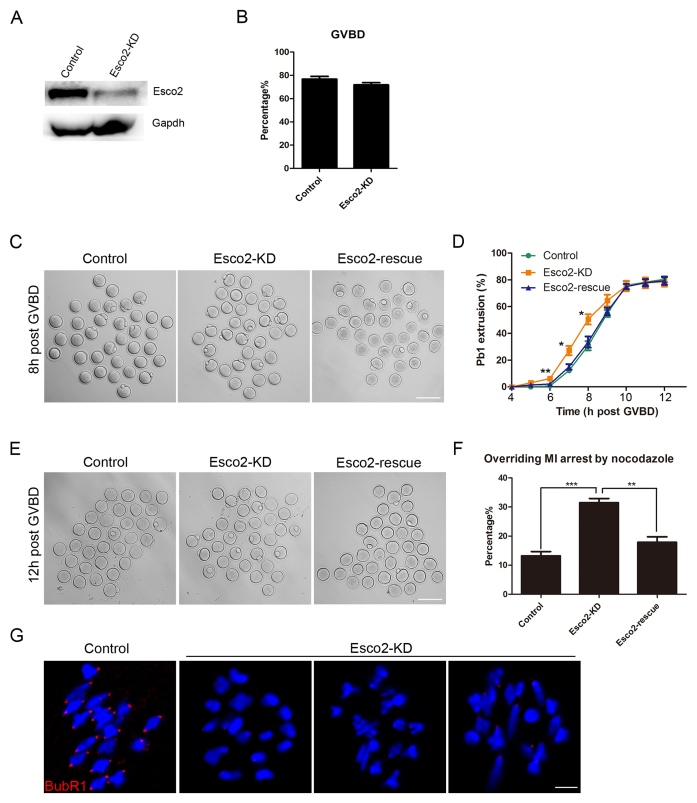
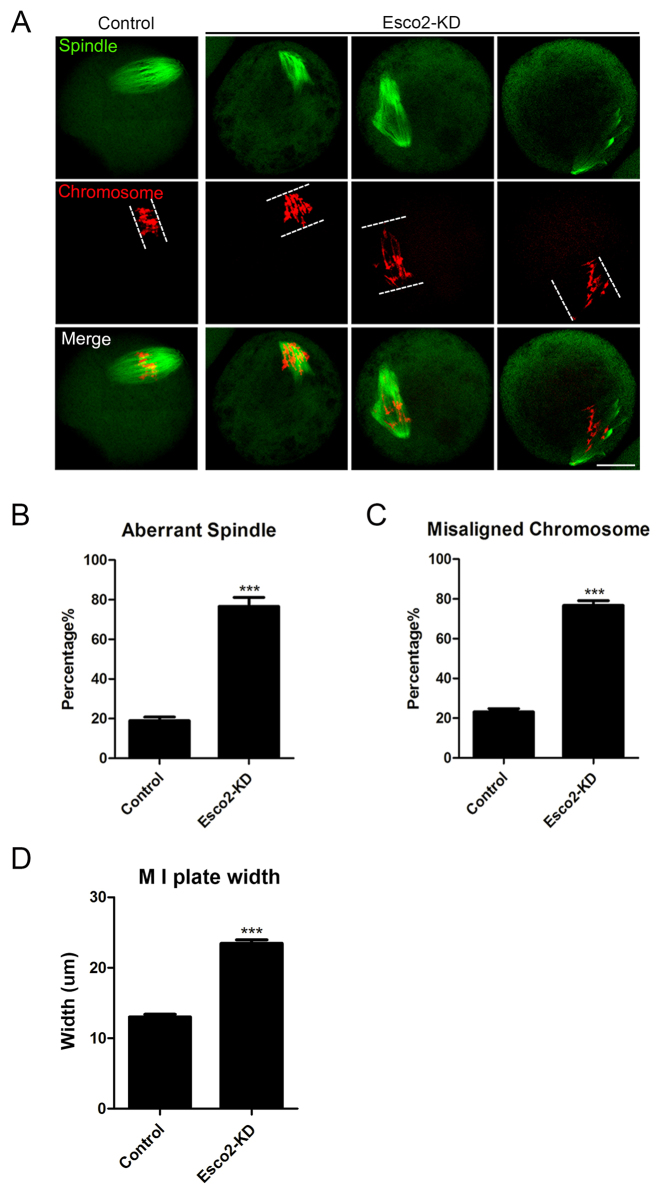
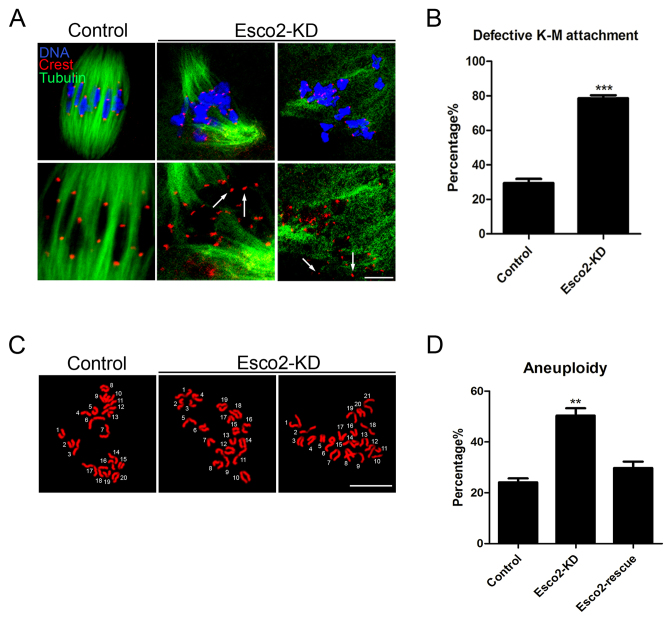
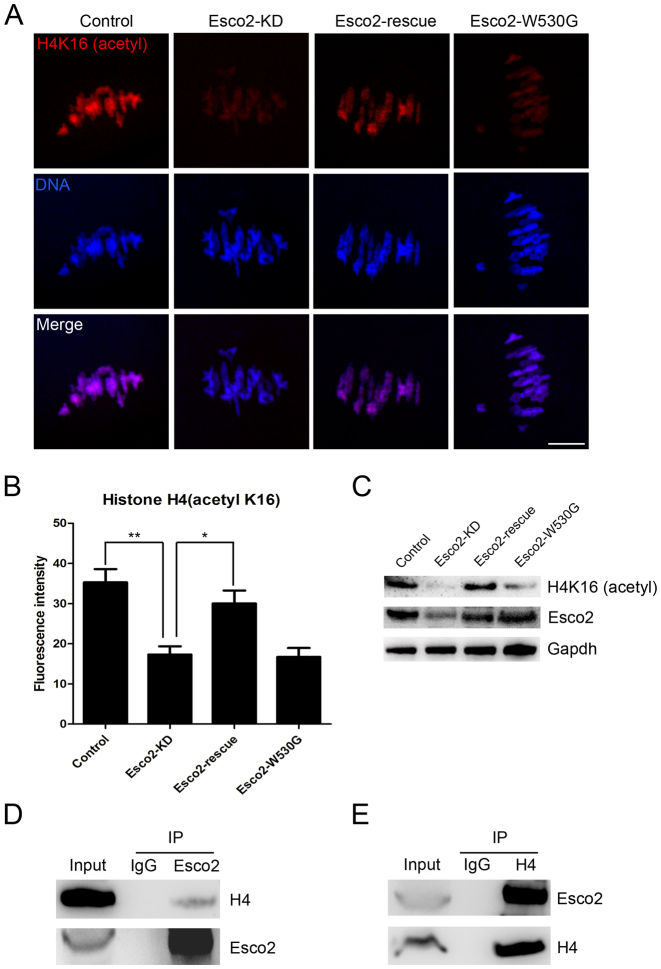
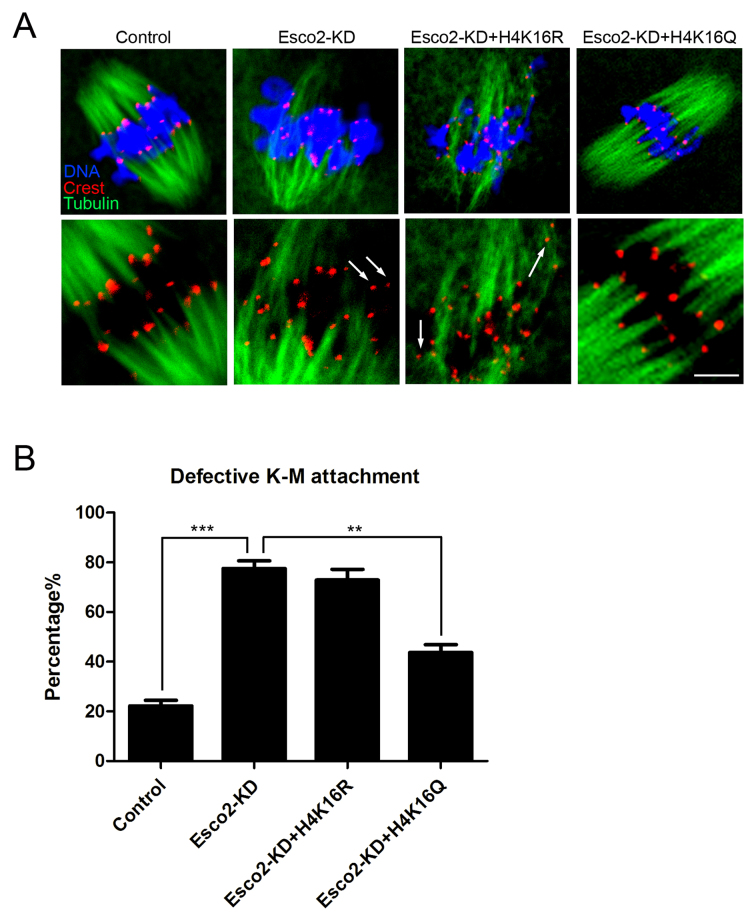
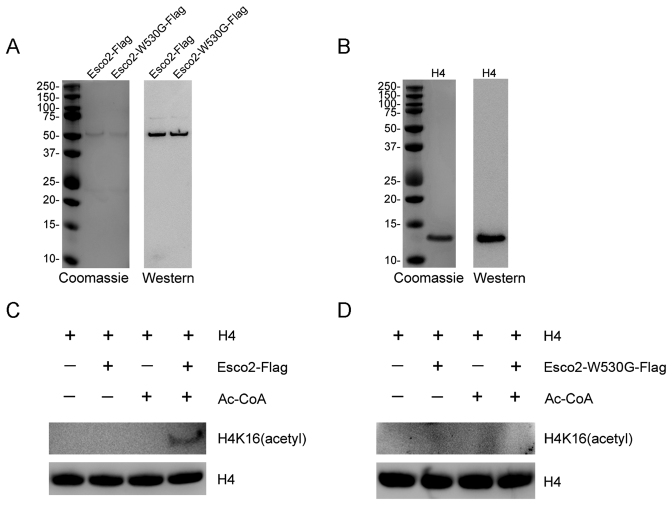
Sister chromatid cohesion, mediated by cohesin complex and established by the acetyltransferases Esco1 and Esco2, is essential for faithful chromosome segregation. Mutations in Esco2 cause Roberts syndrome, a developmental disease characterized by severe prenatal retardation as well as limb and facial abnormalities. However, its exact roles during oocyte meiosis have not clearly defined. Here, we report that Esco2 localizes to the chromosomes during oocyte meiotic maturation. Depletion of Esco2 by morpholino microinjection leads to the precocious polar body extrusion, the escape of metaphase I arrest induced by nocodazole treatment and the loss of BubR1 from kinetochores, indicative of inactivated SAC. Furthermore, depletion of Esco2 causes a severely impaired spindle assembly and chromosome alignment, accompanied by the remarkably elevated incidence of defective kinetochore-microtubule attachments which consequently lead to the generation of aneuploid eggs. Notably, we find that the involvement of Esco2 in SAC and kinetochore functions is mediated by its binding to histone H4 and acetylation of H4K16 both in vivo and in vitro. Thus, our data assign a novel meiotic function to Esco2 beyond its role in the cohesion establishment during mouse oocyte meiosis.
© The Author(s) 2017. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Xiong B., Gerton J.L.. Regulators of the cohesin network. Annu. Rev. Biochem. 2010; 79:131–153. - PubMed
-
- Onn I., Heidinger-Pauli J.M., Guacci V., Unal E., Koshland D.E.. Sister chromatid cohesion: a simple concept with a complex reality. Annu. Rev. Cell Dev. Biol. 2008; 24:105–129. - PubMed
-
- Nasmyth K., Haering C.H.. Cohesin: its roles and mechanisms. Annu. Rev. Genet. 2009; 43:525–558. - PubMed
-
- Saito Y., Zhou H., Kobayashi J.. Chromatin modification and NBS1: their relationship in DNA double-strand break repair. Genes Genet. Syst. 2016; 90:195–208. - PubMed
-
- Rolef Ben-Shahar T., Heeger S., Lehane C., East P., Flynn H., Skehel M., Uhlmann F.. Eco1-dependent cohesin acetylation during establishment of sister chromatid cohesion. Science. 2008; 321:563–566. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
