

Controlled re-activation of epigenetically silenced Tet promoter-driven transgene expression by targeted demethylation
- PMID: 28934472
- PMCID: PMC5766184
- DOI: 10.1093/nar/gkx601
Controlled re-activation of epigenetically silenced Tet promoter-driven transgene expression by targeted demethylation
Abstract
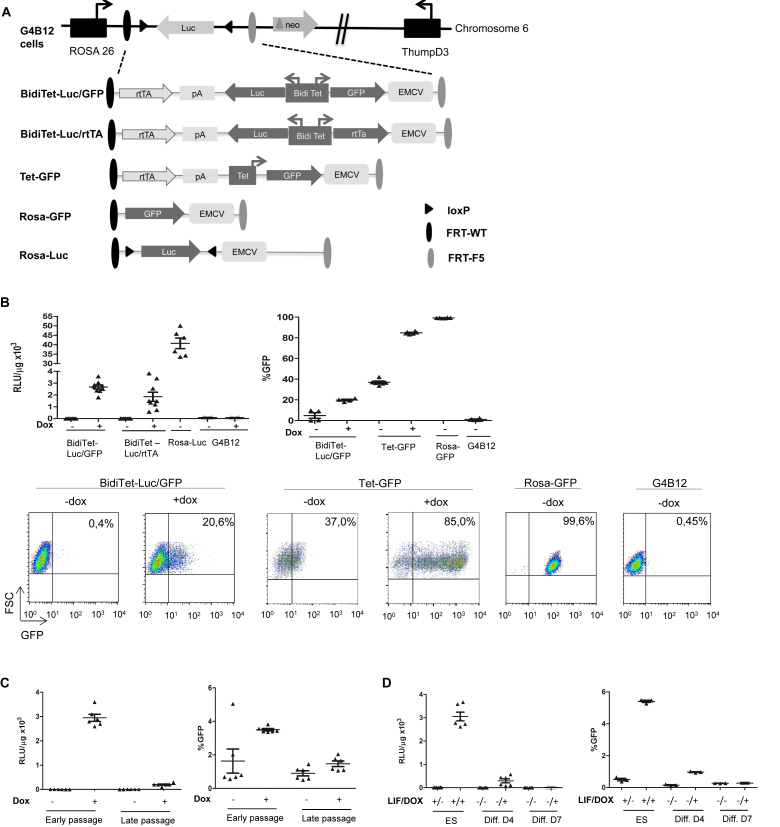
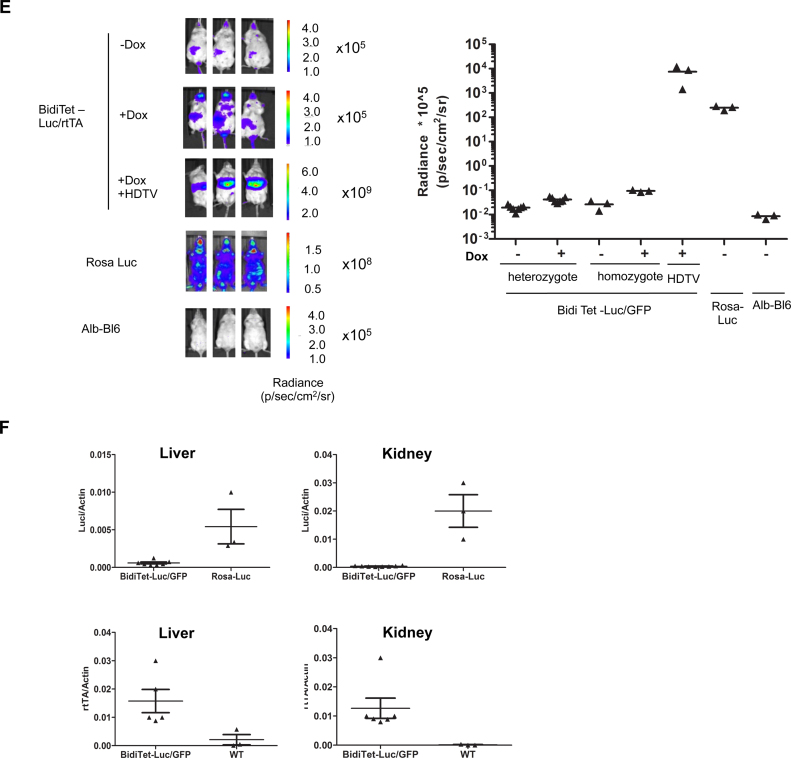
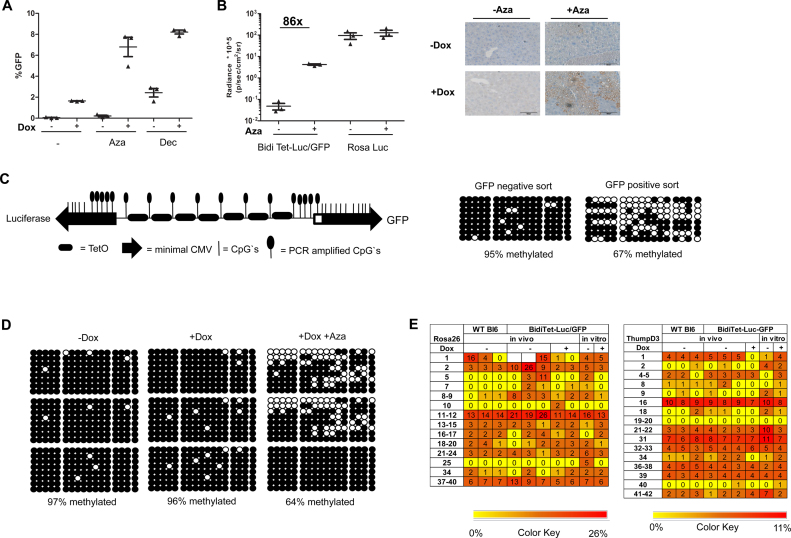
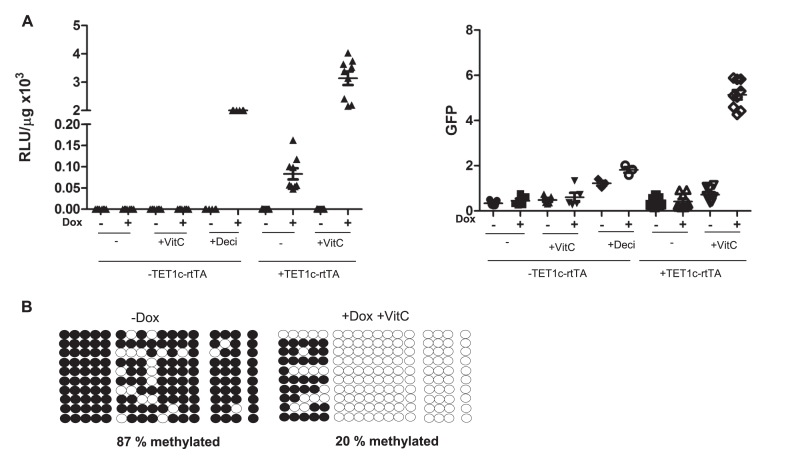
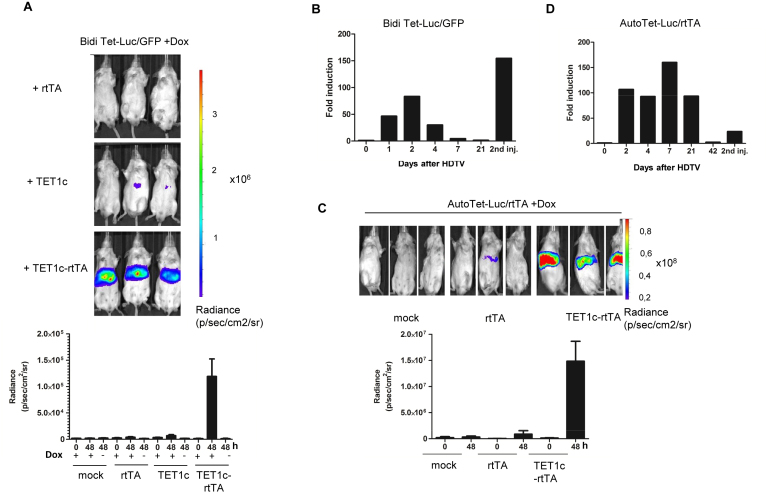
Faithful expression of transgenes in cell cultures and mice is often challenged by locus dependent epigenetic silencing. We investigated silencing of Tet-controlled expression cassettes within the mouse ROSA26 locus. We observed pronounced DNA methylation of the Tet promoter concomitant with loss of expression in mES cells as well as in differentiated cells and transgenic animals. Strikingly, the ROSA26 promoter remains active and methylation free indicating that this silencing mechanism specifically affects the transgene, but does not spread to the host's chromosomal neighborhood. To reactivate Tet cassettes a synthetic fusion protein was constructed and expressed in silenced cells. This protein includes the enzymatic domains of ten eleven translocation methylcytosine dioxygenase 1 (TET-1) as well as the Tet repressor DNA binding domain. Expression of the synthetic fusion protein and Doxycycline treatment allowed targeted demethylation of the Tet promoter in the ROSA26 locus and in another genomic site, rescuing transgene expression in cells and transgenic mice. Thus, inducible, reversible and site-specific epigenetic modulation is a promising strategy for reactivation of silenced transgene expression, independent of the integration site.
© The Author(s) 2017. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Similar articles
-
A Ubiquitous Chromatin Opening Element and DNA Demethylation Facilitate Doxycycline-Controlled Expression during Differentiation and in Transgenic Mice.ACS Synth Biol. 2023 Feb 17;12(2):482-491. doi: 10.1021/acssynbio.2c00450. Epub 2023 Feb 8. ACS Synth Biol. 2023. PMID: 36755406 Free PMC article.
-
Expression of transgenes targeted to the Gt(ROSA)26Sor locus is orientation dependent.PLoS One. 2006 Dec 20;1(1):e4. doi: 10.1371/journal.pone.0000004. PLoS One. 2006. PMID: 17183668 Free PMC article.
-
In Vitro Screen to Identify Silent but Activatable (S/A) Integration Sites for a Tetracycline-Inducible Transgene in Mice.Cold Spring Harb Protoc. 2018 Dec 3;2018(12). doi: 10.1101/pdb.prot092684. Cold Spring Harb Protoc. 2018. PMID: 30510124
-
TET Family of Dioxygenases: Crucial Roles and Underlying Mechanisms.Cytogenet Genome Res. 2015;146(3):171-80. doi: 10.1159/000438853. Epub 2015 Aug 21. Cytogenet Genome Res. 2015. PMID: 26302812 Review.
-
Silencing of transgene expression in mammalian cells by DNA methylation and histone modifications in gene therapy perspective.Biotechnol Genet Eng Rev. 2019 Apr;35(1):1-25. doi: 10.1080/02648725.2018.1551594. Epub 2018 Dec 4. Biotechnol Genet Eng Rev. 2019. PMID: 30514178 Review.
Cited by
-
Programming megakaryocytes to produce engineered platelets for delivering non-native proteins.Commun Biol. 2025 Apr 19;8(1):638. doi: 10.1038/s42003-025-08017-8. Commun Biol. 2025. PMID: 40253534 Free PMC article.
-
Using recombinase-mediated cassette exchange to engineer MIN6 insulin-secreting cells based on a newly identified safe harbor locus.J Diabetes Investig. 2021 Dec;12(12):2129-2140. doi: 10.1111/jdi.13646. Epub 2021 Sep 6. J Diabetes Investig. 2021. PMID: 34382357 Free PMC article.
-
Overcoming the Silencing of Doxycycline-Inducible Promoters in hiPSC-derived Cardiomyocytes.Open Res Eur. 2024 Dec 18;4:266. doi: 10.12688/openreseurope.19024.1. eCollection 2024. Open Res Eur. 2024. PMID: 39926351 Free PMC article.
-
Xist ribonucleoproteins promote female sex-biased autoimmunity.Cell. 2024 Feb 1;187(3):733-749.e16. doi: 10.1016/j.cell.2023.12.037. Cell. 2024. PMID: 38306984 Free PMC article.
-
An Update on Reported Variants in the Skeletal Muscle α-Actin (ACTA1) Gene.Hum Mutat. 2024 Oct 28;2024:6496088. doi: 10.1155/2024/6496088. eCollection 2024. Hum Mutat. 2024. PMID: 40225930 Free PMC article. Review.
References
-
- May T., Butueva M., Bantner S., Markusic D., Seppen J., MacLeod R.A., Weich H., Hauser H., Wirth D.. Synthetic gene regulation circuits for control of cell expansion. Tissue Eng. Part A. 2010; 16:441–452. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
