

The conserved AU dinucleotide at the 5' end of nascent U1 snRNA is optimized for the interaction with nuclear cap-binding-complex
- PMID: 28934473
- PMCID: PMC5766165
- DOI: 10.1093/nar/gkx608
The conserved AU dinucleotide at the 5' end of nascent U1 snRNA is optimized for the interaction with nuclear cap-binding-complex
Abstract
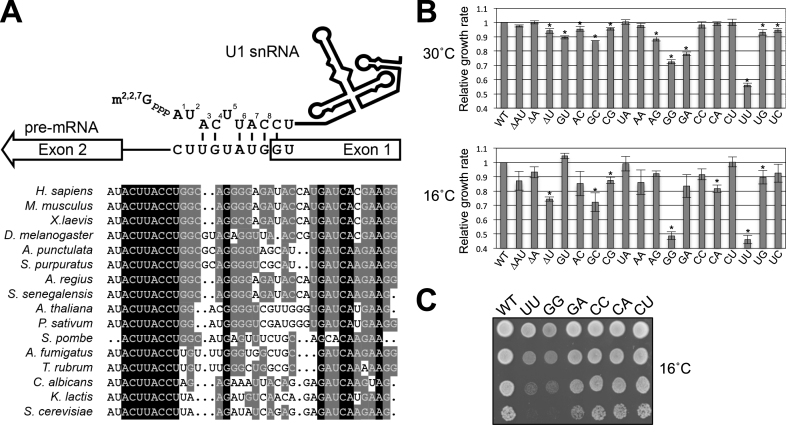
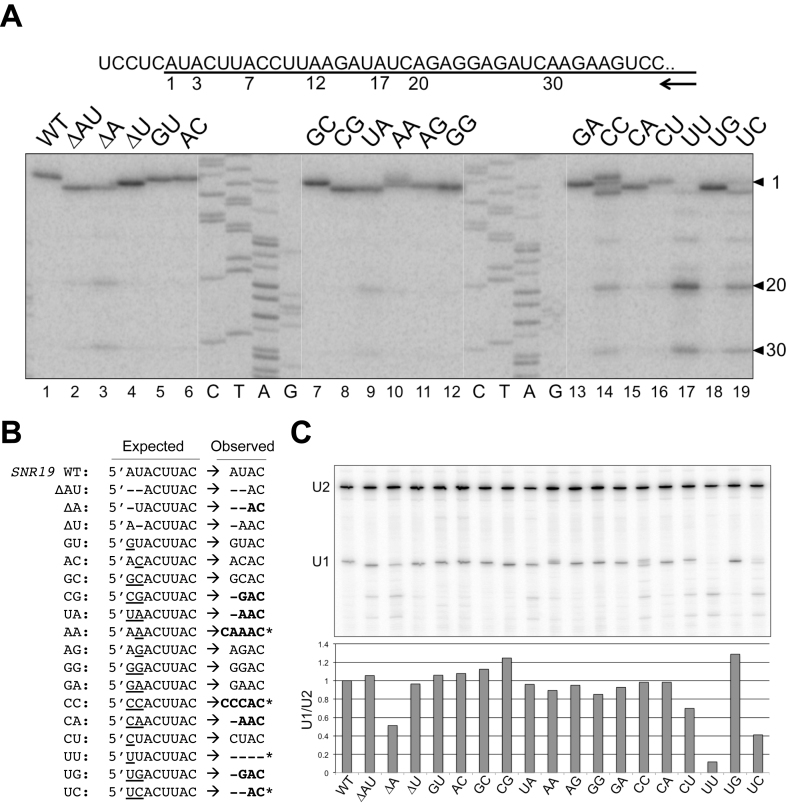
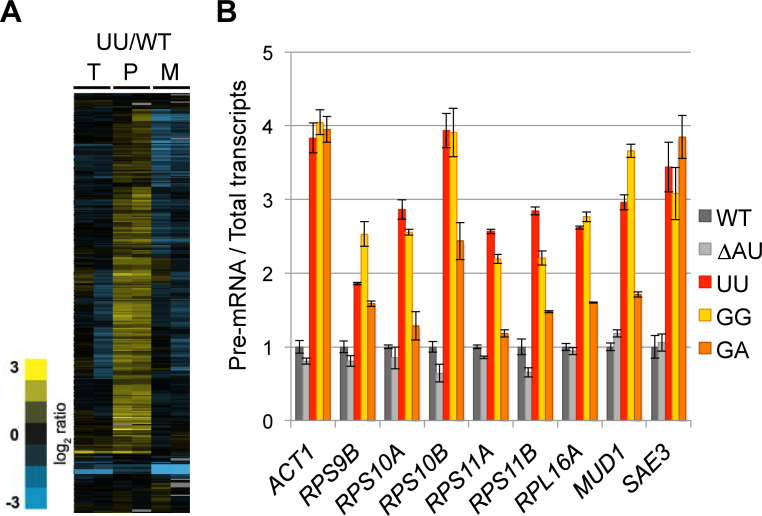
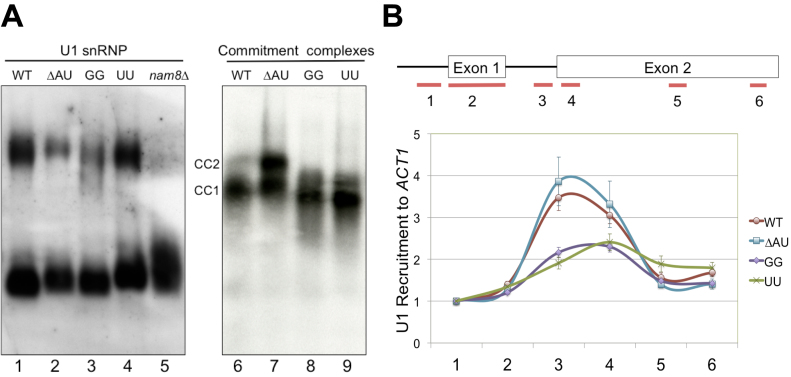
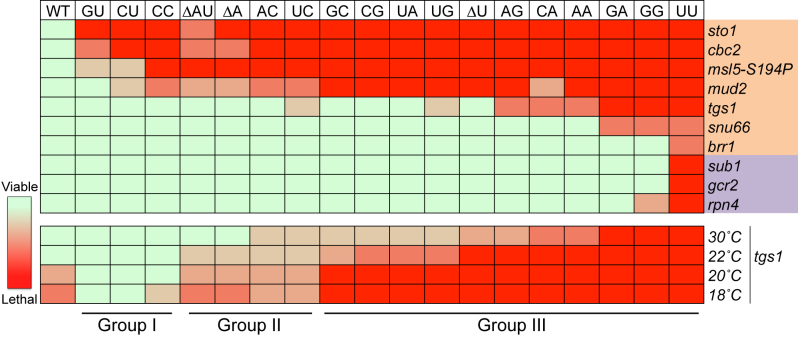
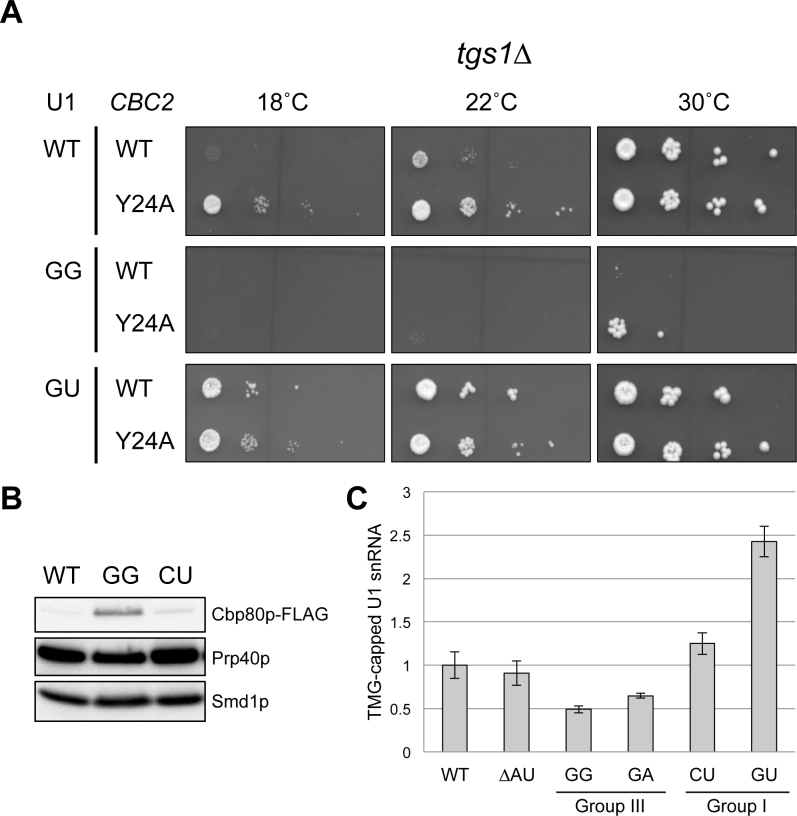
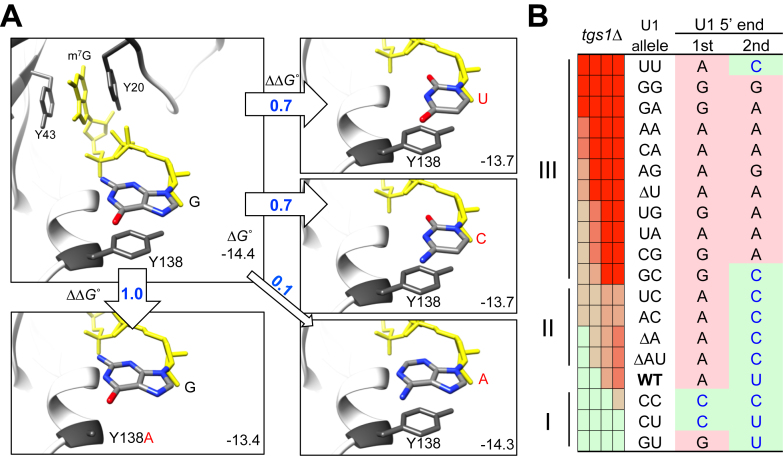
Splicing is initiated by a productive interaction between the pre-mRNA and the U1 snRNP, in which a short RNA duplex is established between the 5' splice site of a pre-mRNA and the 5' end of the U1 snRNA. A long-standing puzzle has been why the AU dincucleotide at the 5'-end of the U1 snRNA is highly conserved, despite the absence of an apparent role in the formation of the duplex. To explore this conundrum, we varied this AU dinucleotide into all possible permutations and analyzed the resulting molecular consequences. This led to the unexpected findings that the AU dinucleotide dictates the optimal binding of cap-binding complex (CBC) to the 5' end of the nascent U1 snRNA, which ultimately influences the utilization of U1 snRNP in splicing. Our data also provide a structural interpretation as to why the AU dinucleotide is conserved during evolution.
© The Author(s) 2017. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Similar articles
-
The U1 snRNP protein U1C recognizes the 5' splice site in the absence of base pairing.Nature. 2002 Sep 5;419(6902):86-90. doi: 10.1038/nature00947. Nature. 2002. PMID: 12214237
-
Yeast U1 snRNP-pre-mRNA complex formation without U1snRNA-pre-mRNA base pairing.RNA. 2001 Jan;7(1):133-42. doi: 10.1017/s1355838201001844. RNA. 2001. PMID: 11214175 Free PMC article.
-
Structure-function analysis of the 5' end of yeast U1 snRNA highlights genetic interactions with the Msl5*Mud2 branchpoint-binding complex and other spliceosome assembly factors.Nucleic Acids Res. 2013 Aug;41(15):7485-500. doi: 10.1093/nar/gkt490. Epub 2013 Jun 10. Nucleic Acids Res. 2013. PMID: 23754852 Free PMC article.
-
[The role of the U1 snRNP complex in the regulation of gene expression: recent reports].Postepy Biochem. 2024 Sep 17;70(3):348-357. doi: 10.18388/pb.2021_566. Print 2024 Sep 30. Postepy Biochem. 2024. PMID: 39365565 Review. Polish.
-
Who's on first? The U1 snRNP-5' splice site interaction and splicing.Trends Biochem Sci. 1991 May;16(5):187-90. doi: 10.1016/0968-0004(91)90073-5. Trends Biochem Sci. 1991. PMID: 1882420 Review.
Cited by
-
Deletions of singular U1 snRNA gene significantly interfere with transcription and 3'-end mRNA formation.PLoS Genet. 2023 Nov 2;19(11):e1011021. doi: 10.1371/journal.pgen.1011021. eCollection 2023 Nov. PLoS Genet. 2023. PMID: 37917726 Free PMC article.
-
The nuclear cap-binding complex as choreographer of gene transcription and pre-mRNA processing.Genes Dev. 2020 Sep 1;34(17-18):1113-1127. doi: 10.1101/gad.339986.120. Genes Dev. 2020. PMID: 32873578 Free PMC article. Review.
-
A novel synthetic-genetic-array-based yeast one-hybrid system for high discovery rate and short processing time.Genome Res. 2019 Aug;29(8):1343-1351. doi: 10.1101/gr.245951.118. Epub 2019 Jun 11. Genome Res. 2019. PMID: 31186303 Free PMC article.
-
Integration of accessibility data from structure probing into RNA-RNA interaction prediction.Bioinformatics. 2019 Aug 15;35(16):2862-2864. doi: 10.1093/bioinformatics/bty1029. Bioinformatics. 2019. PMID: 30590479 Free PMC article.
-
A U1i RNA that Enhances HIV-1 RNA Splicing with an Elongated Recognition Domain Is an Optimal Candidate for Combination HIV-1 Gene Therapy.Mol Ther Nucleic Acids. 2019 Dec 6;18:815-830. doi: 10.1016/j.omtn.2019.10.011. Epub 2019 Oct 18. Mol Ther Nucleic Acids. 2019. PMID: 31734561 Free PMC article.
References
-
- Fabrizio P., Dannenberg J., Dube P., Kastner B., Stark H., Urlaub H., Luhrmann R.. The evolutionarily conserved core design of the catalytic activation step of the yeast spliceosome. Mol. Cell. 2009; 36:593–608. - PubMed
-
- Will C.L., Luhrmann R.. Protein functions in pre-mRNA splicing. Curr. Opin. Cell Biol. 1997; 9:320–328. - PubMed
-
- Chang T.H., Tung L., Yeh F.L., Chen J.H., Chang S.L.. Functions of the DExD/H-box proteins in nuclear pre-mRNA splicing. Biochim. Biophys. Acta. 2013; 1829:764–774. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
