

A germline-limited piggyBac transposase gene is required for precise excision in Tetrahymena genome rearrangement
- PMID: 28934495
- PMCID: PMC5766162
- DOI: 10.1093/nar/gkx652
A germline-limited piggyBac transposase gene is required for precise excision in Tetrahymena genome rearrangement
Abstract
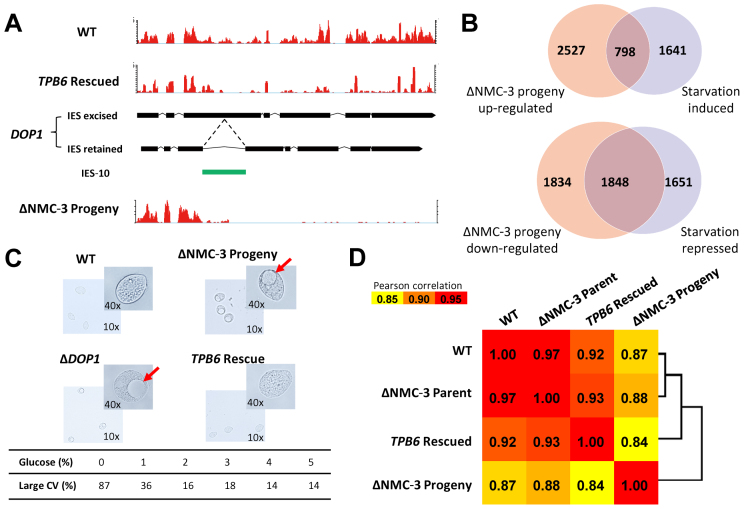
Developmentally programmed genome rearrangement accompanies differentiation of the silent germline micronucleus into the transcriptionally active somatic macronucleus in the ciliated protozoan Tetrahymena thermophila. Internal eliminated sequences (IES) are excised, followed by rejoining of MAC-destined sequences, while fragmentation occurs at conserved chromosome breakage sequences, generating macronuclear chromosomes. Some macronuclear chromosomes, referred to as non-maintained chromosomes (NMC), are lost soon after differentiation. Large NMC contain genes implicated in development-specific roles. One such gene encodes the domesticated piggyBac transposase TPB6, required for heterochromatin-dependent precise excision of IES residing within exons of functionally important genes. These conserved exonic IES determine alternative transcription products in the developing macronucleus; some even contain free-standing genes. Examples of precise loss of some exonic IES in the micronucleus and retention of others in the macronucleus of related species suggest an evolutionary analogy to introns. Our results reveal that germline-limited sequences can encode genes with specific expression patterns and development-related functions, which may be a recurring theme in eukaryotic organisms experiencing programmed genome rearrangement during germline to soma differentiation.
© The Author(s) 2017. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures







Similar articles
-
The piggyBac transposon-derived genes TPB1 and TPB6 mediate essential transposon-like excision during the developmental rearrangement of key genes in Tetrahymena thermophila.Genes Dev. 2016 Dec 15;30(24):2724-2736. doi: 10.1101/gad.290460.116. Genes Dev. 2016. PMID: 28087716 Free PMC article.
-
Developmentally programmed, RNA-directed genome rearrangement in Tetrahymena.Dev Growth Differ. 2012 Jan;54(1):108-19. doi: 10.1111/j.1440-169X.2011.01305.x. Epub 2011 Nov 22. Dev Growth Differ. 2012. PMID: 22103557 Free PMC article. Review.
-
A paradox: rapid evolution rates of germline-limited sequences are associated with conserved patterns of rearrangements in cryptic species of Chilodonella uncinata (Protista, Ciliophora).Sci China Life Sci. 2018 Sep;61(9):1071-1078. doi: 10.1007/s11427-018-9333-1. Epub 2018 Jul 19. Sci China Life Sci. 2018. PMID: 30069672
-
Macronuclear genome sequence of the ciliate Tetrahymena thermophila, a model eukaryote.PLoS Biol. 2006 Sep;4(9):e286. doi: 10.1371/journal.pbio.0040286. PLoS Biol. 2006. PMID: 16933976 Free PMC article.
-
Small RNAs in genome rearrangement in Tetrahymena.Curr Opin Genet Dev. 2004 Apr;14(2):181-7. doi: 10.1016/j.gde.2004.01.004. Curr Opin Genet Dev. 2004. PMID: 15196465 Review.
Cited by
-
Programmed DNA elimination: silencing genes and repetitive sequences in somatic cells.Biochem Soc Trans. 2021 Nov 1;49(5):1891-1903. doi: 10.1042/BST20190951. Biochem Soc Trans. 2021. PMID: 34665225 Free PMC article. Review.
-
Programmed genome rearrangements in ciliates.Cell Mol Life Sci. 2020 Nov;77(22):4615-4629. doi: 10.1007/s00018-020-03555-2. Epub 2020 May 27. Cell Mol Life Sci. 2020. PMID: 32462406 Free PMC article. Review.
-
Meiosis and beyond - understanding the mechanistic and evolutionary processes shaping the germline genome.Biol Rev Camb Philos Soc. 2021 Jun;96(3):822-841. doi: 10.1111/brv.12680. Epub 2021 Jan 1. Biol Rev Camb Philos Soc. 2021. PMID: 33615674 Free PMC article. Review.
-
Structural insights into the evolution of the RAG recombinase.Nat Rev Immunol. 2022 Jun;22(6):353-370. doi: 10.1038/s41577-021-00628-6. Epub 2021 Oct 21. Nat Rev Immunol. 2022. PMID: 34675378 Review.
-
A Polycomb repressive complex is required for RNAi-mediated heterochromatin formation and dynamic distribution of nuclear bodies.Nucleic Acids Res. 2021 Jun 4;49(10):5407-5425. doi: 10.1093/nar/gkaa1262. Nucleic Acids Res. 2021. PMID: 33412588 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
