

Suppression of ERK signalling abolishes primitive endoderm formation but does not promote pluripotency in rabbit embryo
- PMID: 28935706
- PMCID: PMC5675450
- DOI: 10.1242/dev.156406
Suppression of ERK signalling abolishes primitive endoderm formation but does not promote pluripotency in rabbit embryo
Abstract
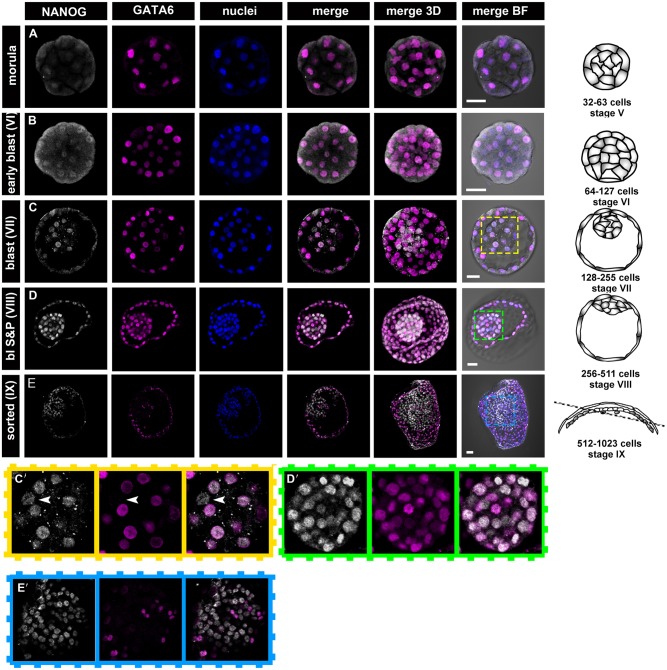
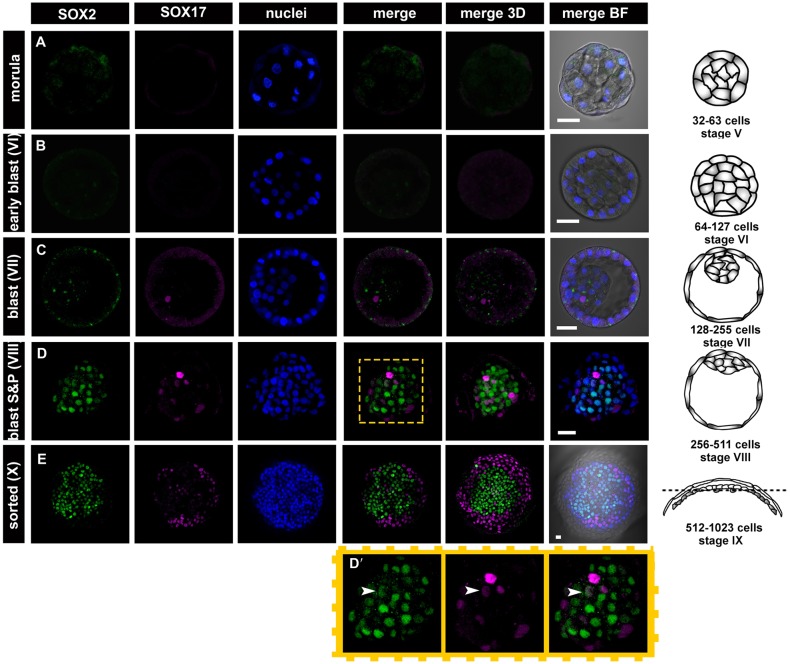
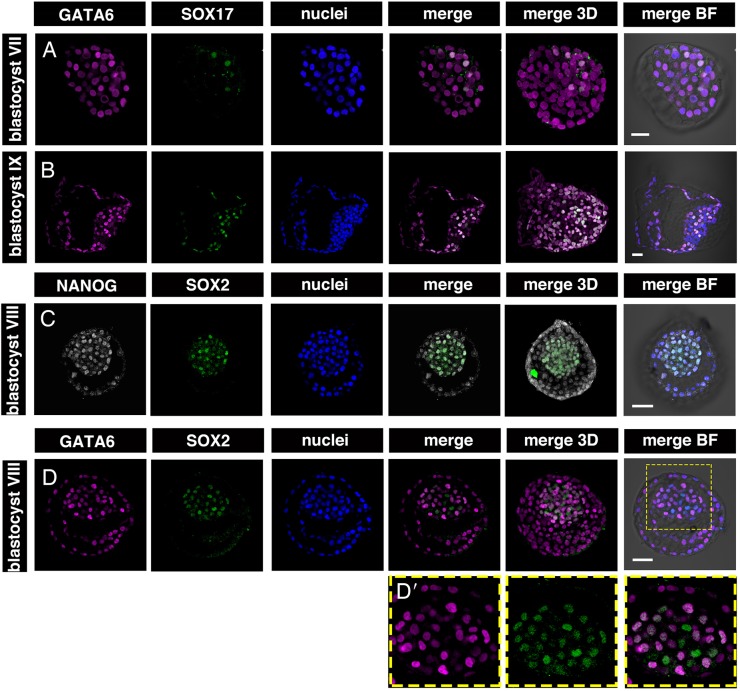
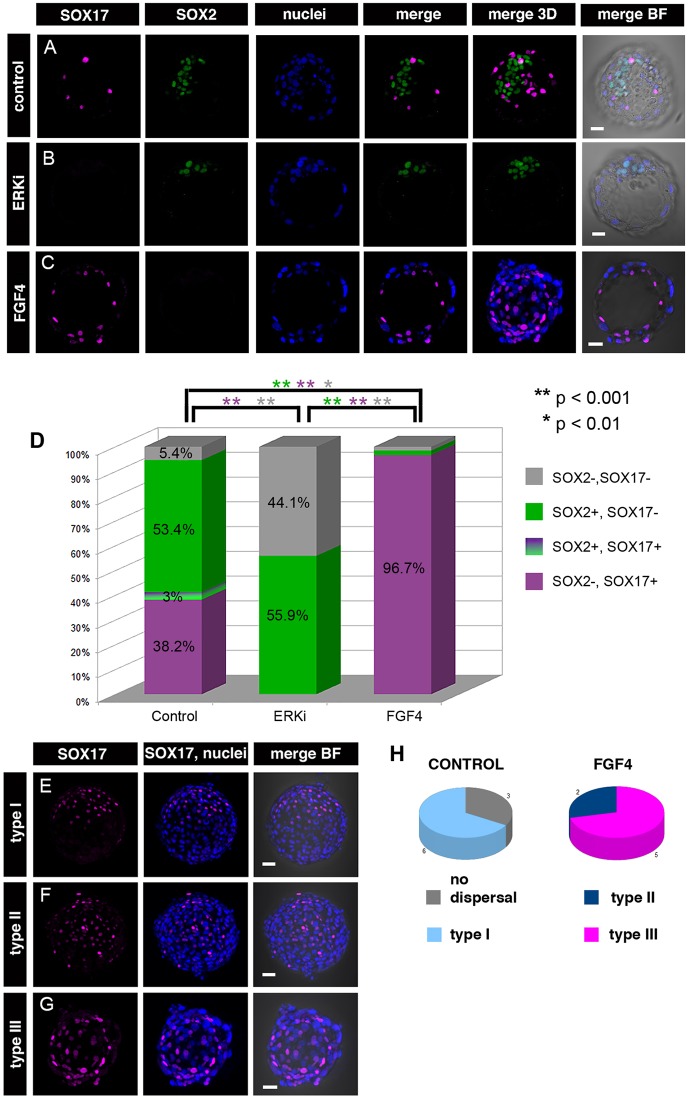
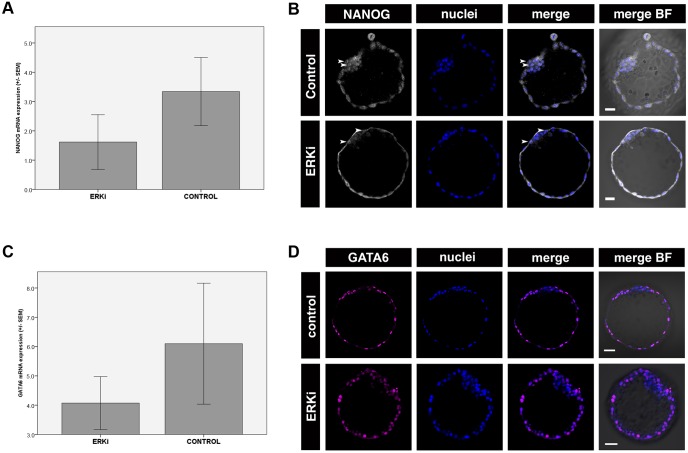
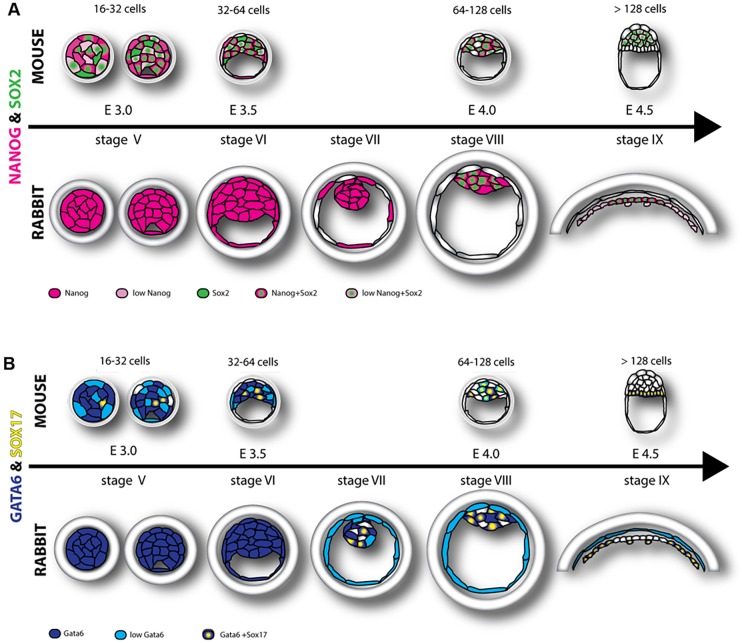
Formation of epiblast (EPI) - the founder line of all embryonic lineages - and extra-embryonic supportive tissues is one of the key events in mammalian development. The prevailing model of early mammalian development is based almost exclusively on the mouse. Here, we provide a comprehensive, stage-by-stage analysis of EPI and extra-embryonic primitive endoderm (PrE) formation during preimplantation development of the rabbit. Although we observed that rabbit embryos have several features in common with mouse embryos, including a stage-related initiation of lineage specification, our results demonstrate the existence of some key differences in lineage specification among mammals. Contrary to the current view, our data suggest that reciprocal repression of GATA6 and NANOG is not fundamental for the initial stages of PrE versus EPI specification in mammals. Furthermore, our results provide insight into the observed discrepancies relating to the role of FGF/ERK signalling in PrE versus EPI specification between mouse and other mammals.
Keywords: Blastocyst; Epiblast; FGF; Primitive endoderm; Rabbit.
© 2017. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
References
-
- Acampora D., Omodei D., Petrosino G., Garofalo A., Savarese M., Nigro V., Di Giovannantonio L. G., Mercadante V. and Simeone A. (2016). Loss of the Otx2-binding site in the Nanog promoter affects the integrity of embryonic stem cell subtypes and specification of inner cell mass-derived epiblast. Cell Rep. 15, 2651-2664. 10.1016/j.celrep.2016.05.041 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
