

Neural-specific deletion of the focal adhesion adaptor protein paxillin slows migration speed and delays cortical layer formation
- PMID: 28935710
- PMCID: PMC5702069
- DOI: 10.1242/dev.147934
Neural-specific deletion of the focal adhesion adaptor protein paxillin slows migration speed and delays cortical layer formation
Abstract
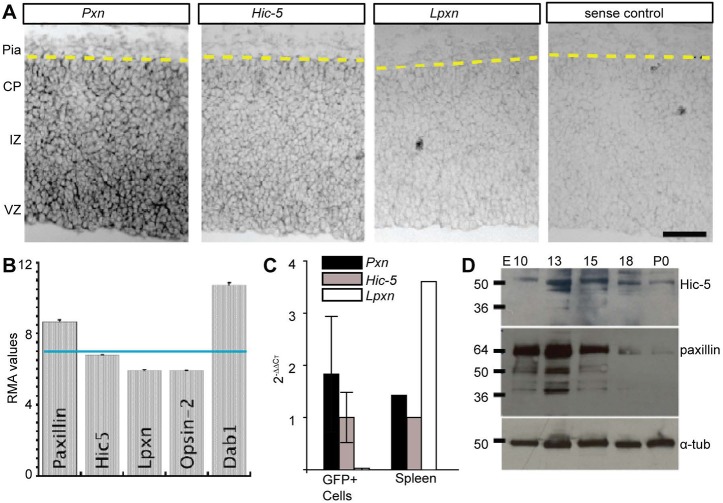
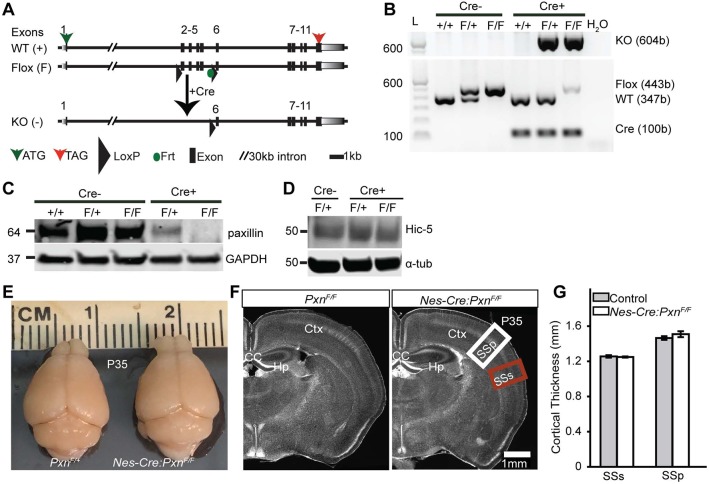
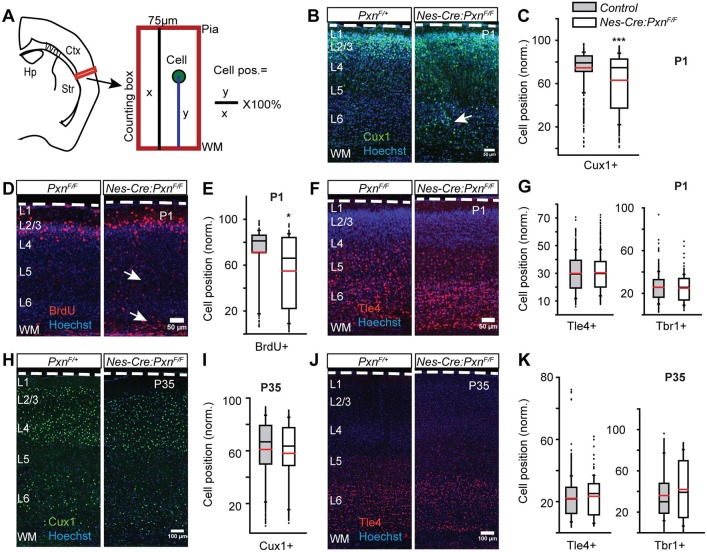
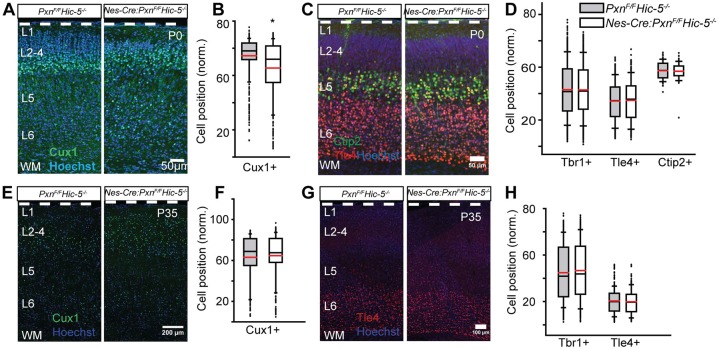
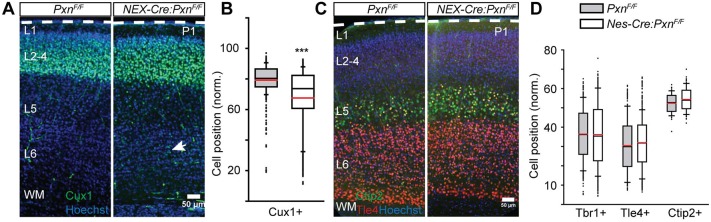
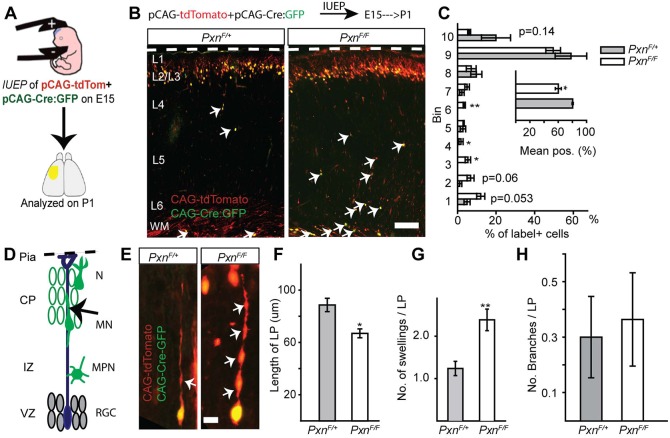
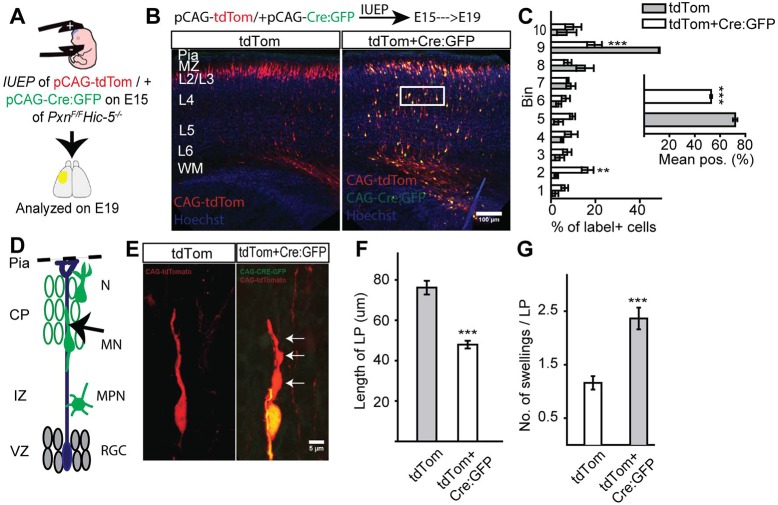
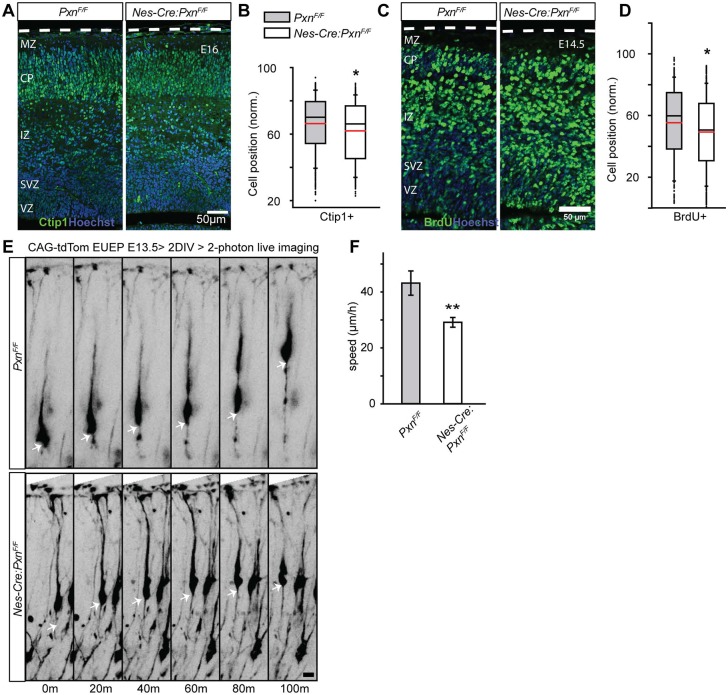
Paxillin and Hic-5 are homologous focal adhesion adaptor proteins that coordinate cytoskeletal rearrangements in response to integrin signaling, but their role(s) in cortical development are unknown. Here, we find that Hic-5-deficient mice are postnatal viable with normal cortical layering. Mice with a neural-specific deletion of paxillin are also postnatal viable, but show evidence of a cortical neuron migration delay that is evident pre- and perinatally, but is not detected at postnatal day 35 (P35). This phenotype is not modified by Hic-5 deficiency (double knockout). Specific deletion of paxillin in postmitotic neurons using Nex-Cre-mediated recombination as well as in utero electroporation of a Cre-expression construct identified a cell-autonomous requirement for paxillin in migrating neurons. Paxillin-deficient neurons have shorter leading processes that exhibited multiple swellings in comparison with control. Multiphoton imaging revealed that paxillin-deficient neurons migrate ∼30% slower than control neurons. This phenotype is similar to that produced by deletion of focal adhesion kinase (FAK), a signaling partner of paxillin, and suggests that paxillin and FAK function cell-autonomously to control migrating neuron morphology and speed during cortical development.
Keywords: Cortical development; Developmental delay; Glial guided migration; Leading process.
© 2017. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
Similar articles
-
Delayed cortical development in mice with a neural specific deletion of β1 integrin.Front Neurosci. 2023 May 12;17:1158419. doi: 10.3389/fnins.2023.1158419. eCollection 2023. Front Neurosci. 2023. PMID: 37250402 Free PMC article.
-
Paxillin and Hic-5 interaction with vinculin is differentially regulated by Rac1 and RhoA.PLoS One. 2012;7(5):e37990. doi: 10.1371/journal.pone.0037990. Epub 2012 May 22. PLoS One. 2012. PMID: 22629471 Free PMC article.
-
A comparative analysis of paxillin and Hic-5 proximity interactomes.Cytoskeleton (Hoboken). 2025 Jan;82(1-2):12-31. doi: 10.1002/cm.21878. Epub 2024 May 27. Cytoskeleton (Hoboken). 2025. PMID: 38801098
-
Paxillin and Kindlin: Research Progress and Biological Functions.Biomolecules. 2025 Jan 24;15(2):173. doi: 10.3390/biom15020173. Biomolecules. 2025. PMID: 40001476 Free PMC article. Review.
-
Paxillin: a crossroad in pathological cell migration.J Hematol Oncol. 2017 Feb 18;10(1):50. doi: 10.1186/s13045-017-0418-y. J Hematol Oncol. 2017. PMID: 28214467 Free PMC article. Review.
Cited by
-
A kindlin-3-leupaxin-paxillin signaling pathway regulates podosome stability.J Cell Biol. 2019 Oct 7;218(10):3436-3454. doi: 10.1083/jcb.201903109. Epub 2019 Sep 19. J Cell Biol. 2019. PMID: 31537712 Free PMC article.
-
Linking Cell Polarity to Cortical Development and Malformations.Front Cell Neurosci. 2019 Jun 4;13:244. doi: 10.3389/fncel.2019.00244. eCollection 2019. Front Cell Neurosci. 2019. PMID: 31213986 Free PMC article. Review.
-
Alveolar epithelial paxillin in postnatal lung alveolar development.Biol Open. 2025 Mar 15;14(3):bio061939. doi: 10.1242/bio.061939. Epub 2025 Mar 20. Biol Open. 2025. PMID: 39991922 Free PMC article.
-
Focal adhesion kinase: from biological functions to therapeutic strategies.Exp Hematol Oncol. 2023 Sep 25;12(1):83. doi: 10.1186/s40164-023-00446-7. Exp Hematol Oncol. 2023. PMID: 37749625 Free PMC article. Review.
-
Ethanol Exposure Transiently Elevates but Persistently Inhibits Tyrosine Kinase Activity and Impairs the Growth of the Nascent Apical Dendrite.Mol Neurobiol. 2019 Aug;56(8):5749-5762. doi: 10.1007/s12035-019-1473-x. Epub 2019 Jan 23. Mol Neurobiol. 2019. PMID: 30674037 Free PMC article.
References
-
- Beggs H. E., Schahin-Reed D., Zang K., Goebbels S., Nave K.-A., Gorski J., Jones K. R., Sretavan D. and Reichardt L. F. (2003). FAK deficiency in cells contributing to the basal lamina results in cortical abnormalities resembling congenital muscular dystrophies. Neuron 40, 501-514. 10.1016/S0896-6273(03)00666-4 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
