

Salinomycin kills cancer stem cells by sequestering iron in lysosomes
- PMID: 28937680
- PMCID: PMC5890907
- DOI: 10.1038/nchem.2778
Salinomycin kills cancer stem cells by sequestering iron in lysosomes
Abstract
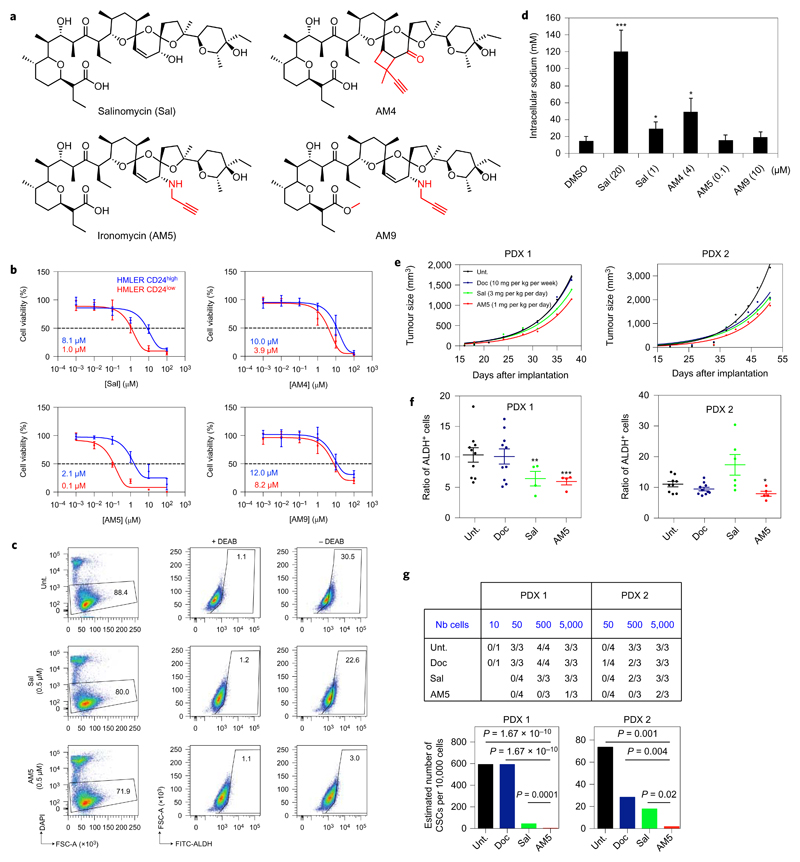
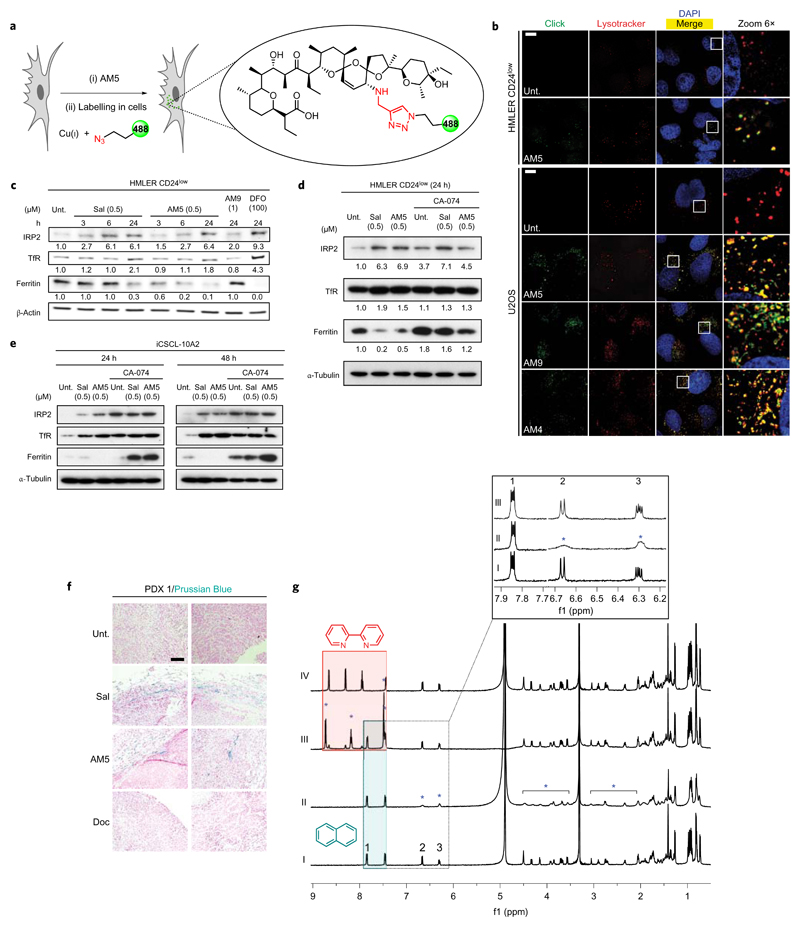
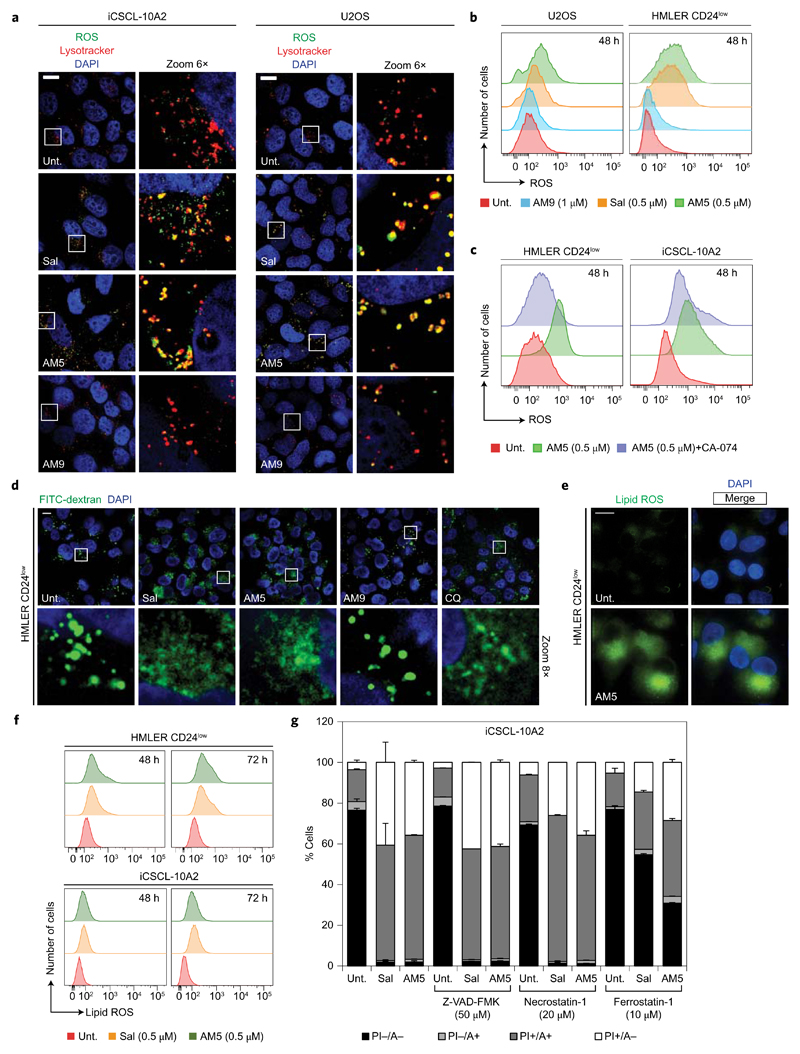
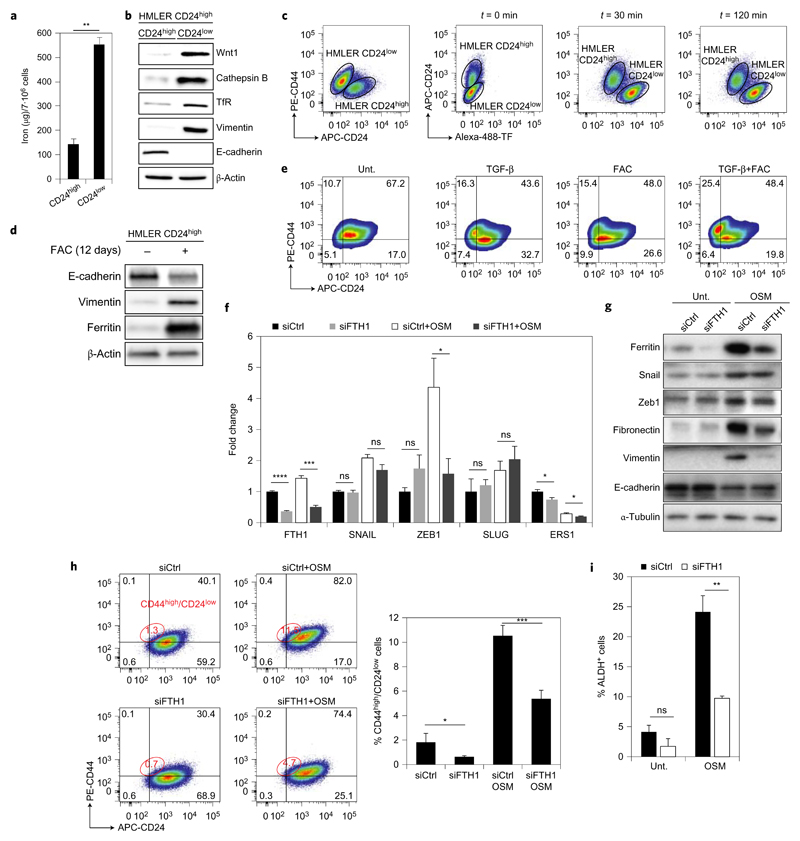
Cancer stem cells (CSCs) represent a subset of cells within tumours that exhibit self-renewal properties and the capacity to seed tumours. CSCs are typically refractory to conventional treatments and have been associated to metastasis and relapse. Salinomycin operates as a selective agent against CSCs through mechanisms that remain elusive. Here, we provide evidence that a synthetic derivative of salinomycin, which we named ironomycin (AM5), exhibits a more potent and selective activity against breast CSCs in vitro and in vivo, by accumulating and sequestering iron in lysosomes. In response to the ensuing cytoplasmic depletion of iron, cells triggered the degradation of ferritin in lysosomes, leading to further iron loading in this organelle. Iron-mediated production of reactive oxygen species promoted lysosomal membrane permeabilization, activating a cell death pathway consistent with ferroptosis. These findings reveal the prevalence of iron homeostasis in breast CSCs, pointing towards iron and iron-mediated processes as potential targets against these cells.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
