

A vitronectin-derived peptide reverses ovariectomy-induced bone loss via regulation of osteoblast and osteoclast differentiation
- PMID: 28937683
- PMCID: PMC5762842
- DOI: 10.1038/cdd.2017.153
A vitronectin-derived peptide reverses ovariectomy-induced bone loss via regulation of osteoblast and osteoclast differentiation
Abstract
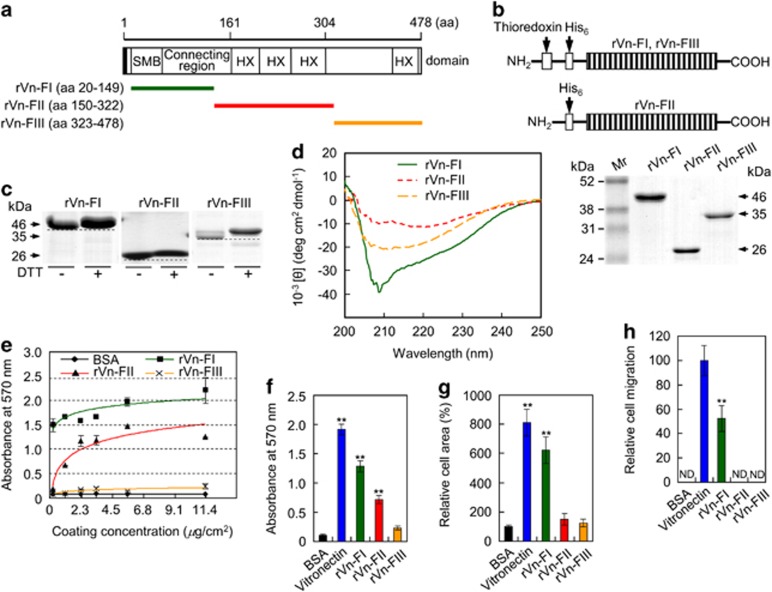
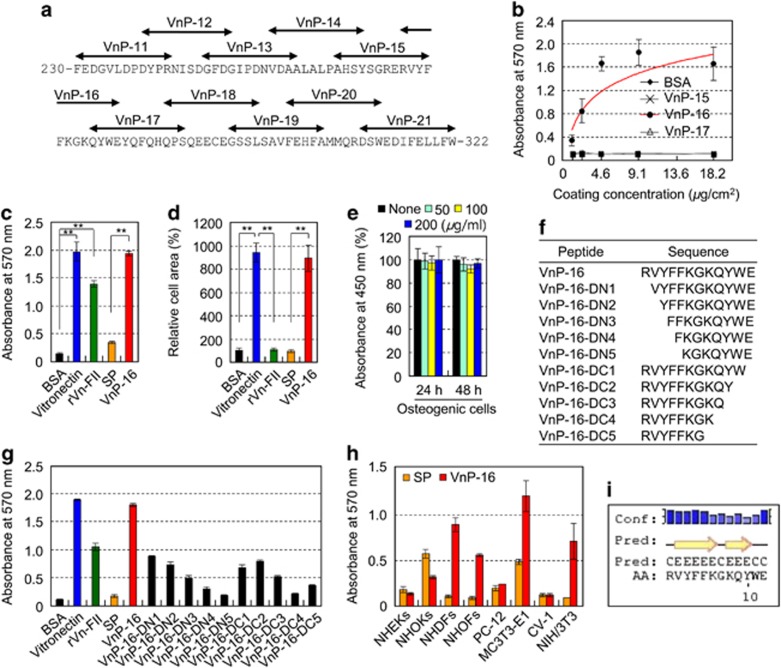
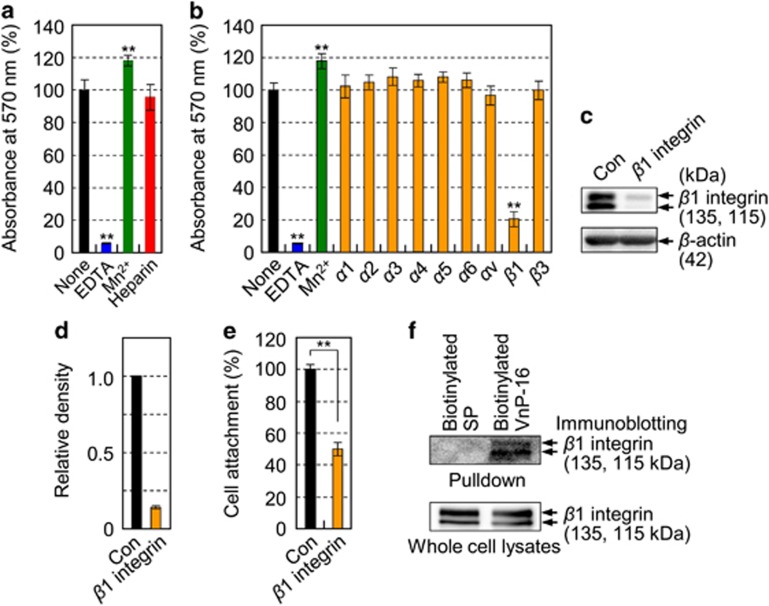
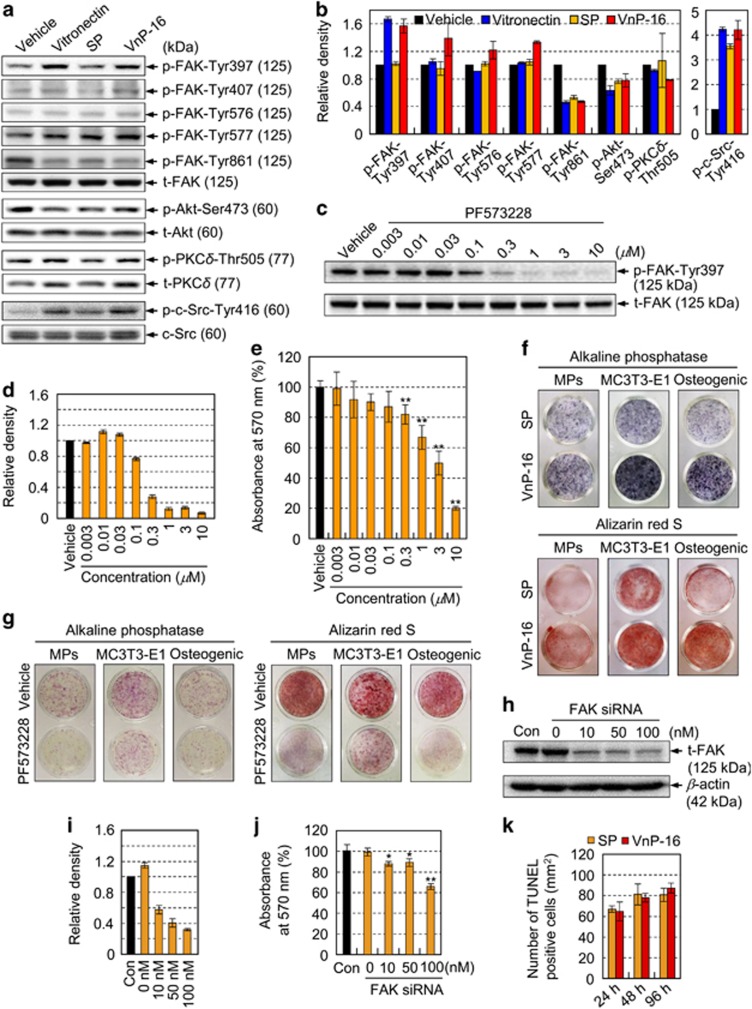
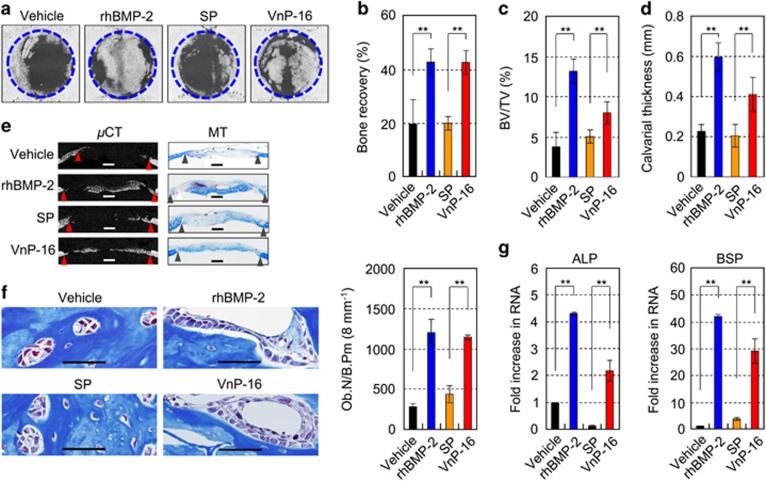
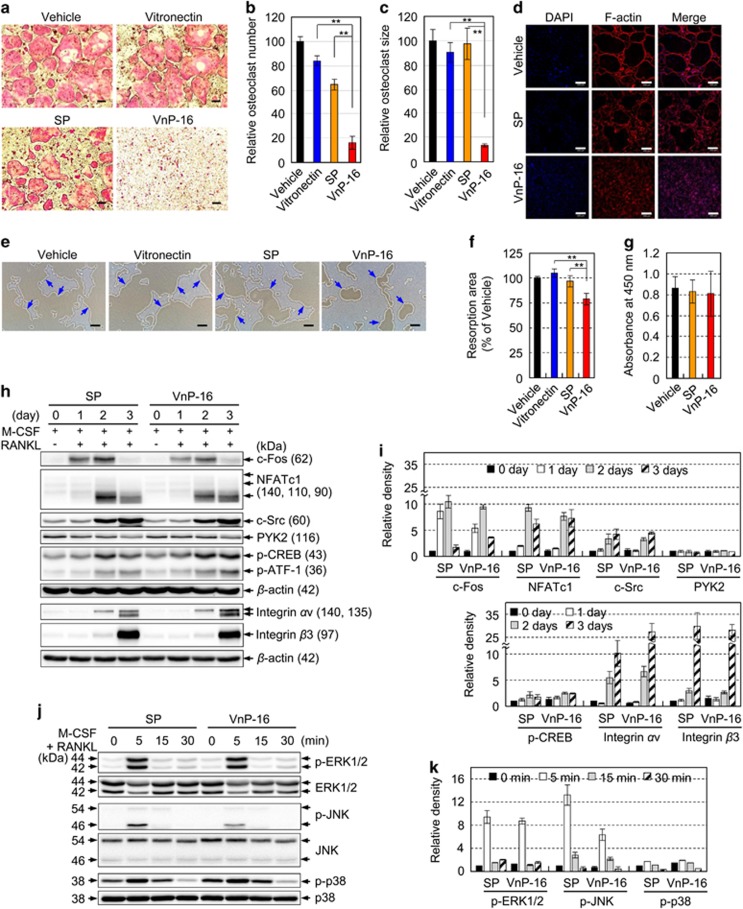
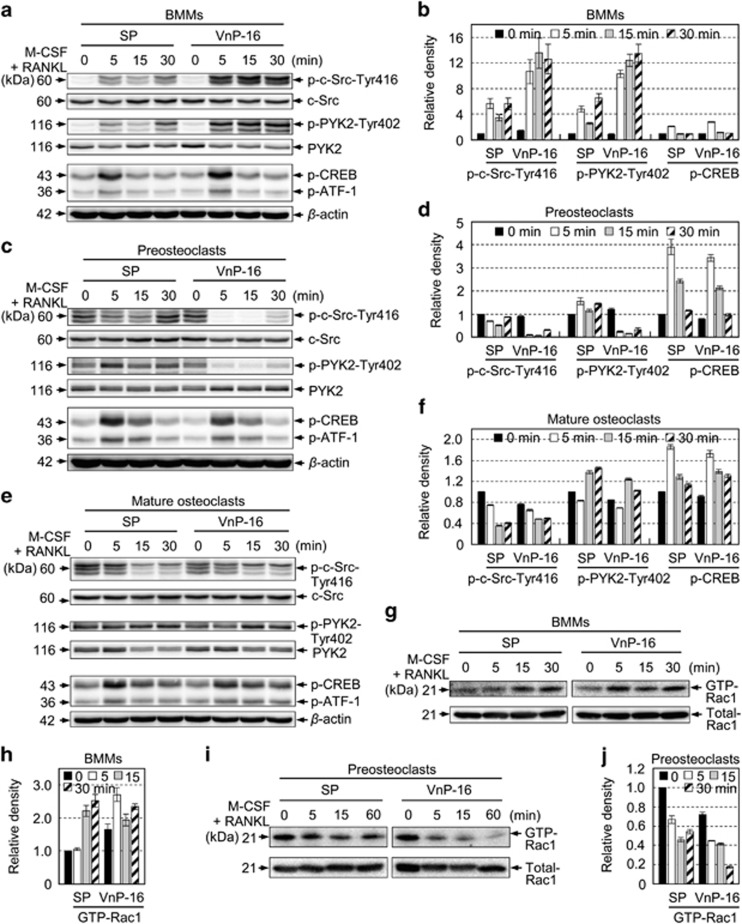
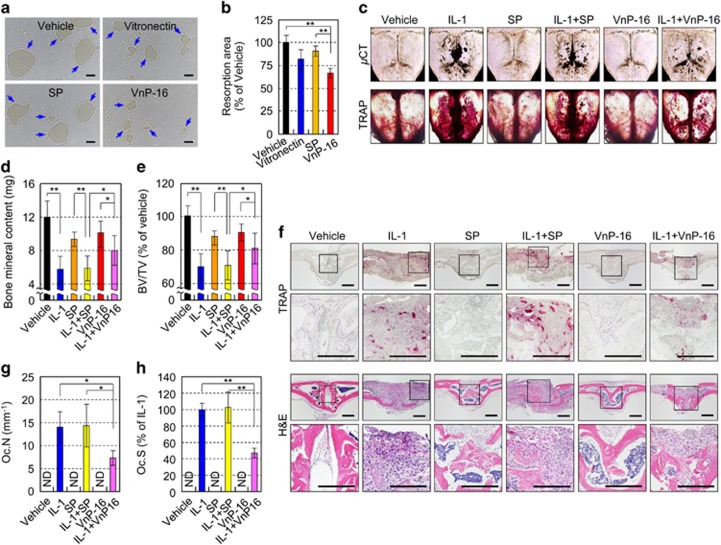
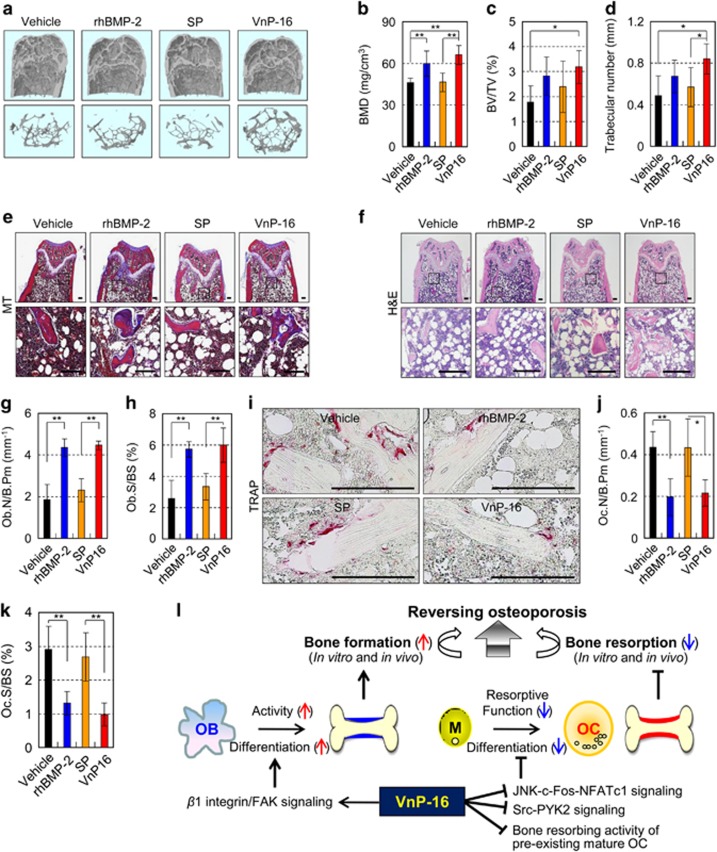
Osteoporosis affects millions of people worldwide by promoting bone resorption and impairing bone formation. Bisphosphonates, commonly used agents to treat osteoporosis, cannot reverse the substantial bone loss that has already occurred by the time of diagnosis. Moreover, their undesirable side-effects, including osteonecrosis of the jaw, have been reported. Here, we demonstrated that a new bioactive core vitronectin-derived peptide (VnP-16) promoted bone formation by accelerating osteoblast differentiation and activity through direct interaction with β1 integrin followed by FAK activation. Concomitantly, VnP-16 inhibited bone resorption by restraining JNK-c-Fos-NFATc1-induced osteoclast differentiation and αvβ3 integrin-c-Src-PYK2-mediated resorptive function. Moreover, VnP-16 decreased the bone resorbing activity of pre-existing mature osteoclasts without changing their survival rate. Furthermore, VnP-16 had a strong anabolic effect on bone regeneration by stimulating osteoblast differentiation and increasing osteoblast number, and significantly alleviated proinflammatory cytokine-induced bone resorption by restraining osteoclast differentiation and function in murine models. Moreover, VnP-16 could reverse ovariectomy-induced bone loss by both inhibiting bone resorption and promoting bone formation. Given its dual role in promoting bone formation and inhibiting bone resorption, our results suggest that VnP-16 could be an attractive therapeutic agent for treating osteoporosis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Schieker M, Mutschler W. Bridging posttraumatic bony defects. Established and new methods. Unfallchirurg 2006; 109: 715–732. - PubMed
-
- Yaszemski MJ, Payne RG, Hayes WC, Langer R, Mikos AG. Evolution of bone transplantation: molecular, cellular and tissue strategies to engineer human bone. Biomaterials 1996; 17: 175–185. - PubMed
-
- Wiesmann HP, Joos U, Meyer U. Biological and biophysical principles in extracorporal bone tissue engineering. Part II. Int J Oral Maxillofac Surg 2004; 33: 523–530. - PubMed
-
- Tanaka S, Nakamura K, Takahasi N, Suda T. Role of RANKL in physiological and pathological bone resorption and therapeutics targeting the RANKL-RANK signaling system. Immunol Rev 2005; 208: 30–49. - PubMed
-
- Broadhead ML, Clark JC, Dass CR, Choong PF, Myers DE. Therapeutic targeting of osteoclast function and pathways. Expert Opin Ther Targets 2011; 15: 169–181. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
