

Regional endothermy as a trigger for gigantism in some extinct macropredatory sharks
- PMID: 28938002
- PMCID: PMC5609766
- DOI: 10.1371/journal.pone.0185185
Regional endothermy as a trigger for gigantism in some extinct macropredatory sharks
Abstract
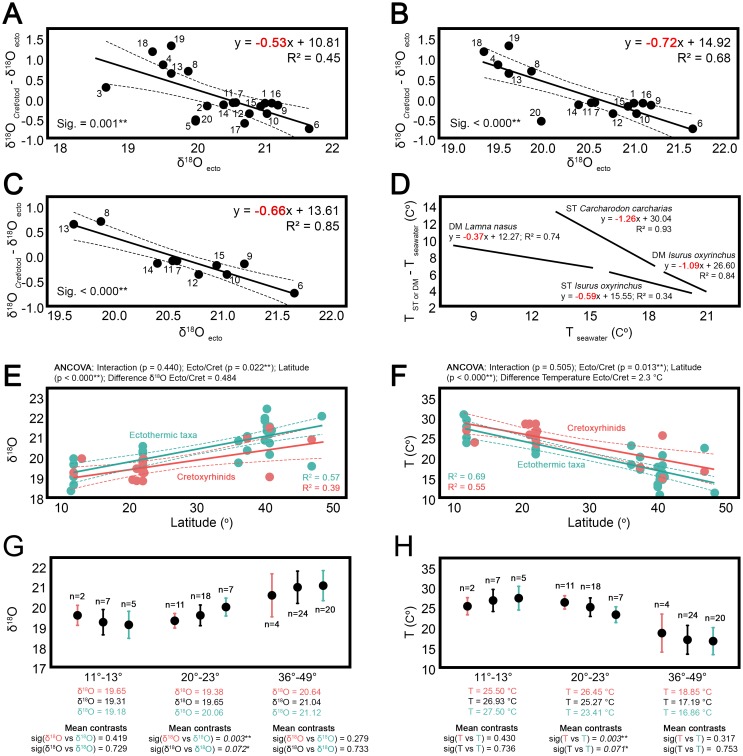
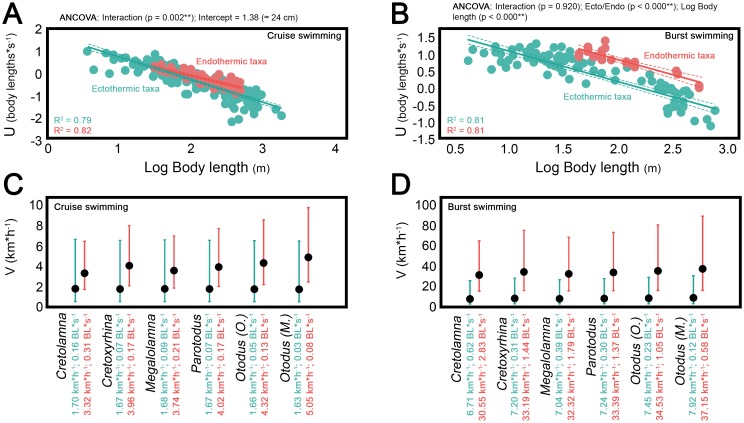
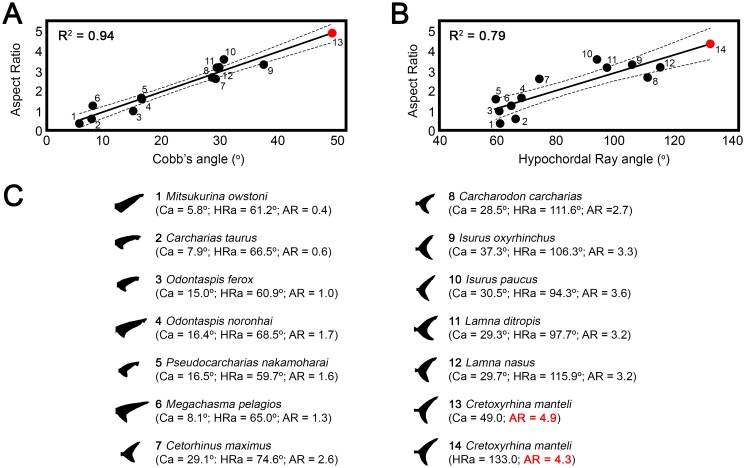
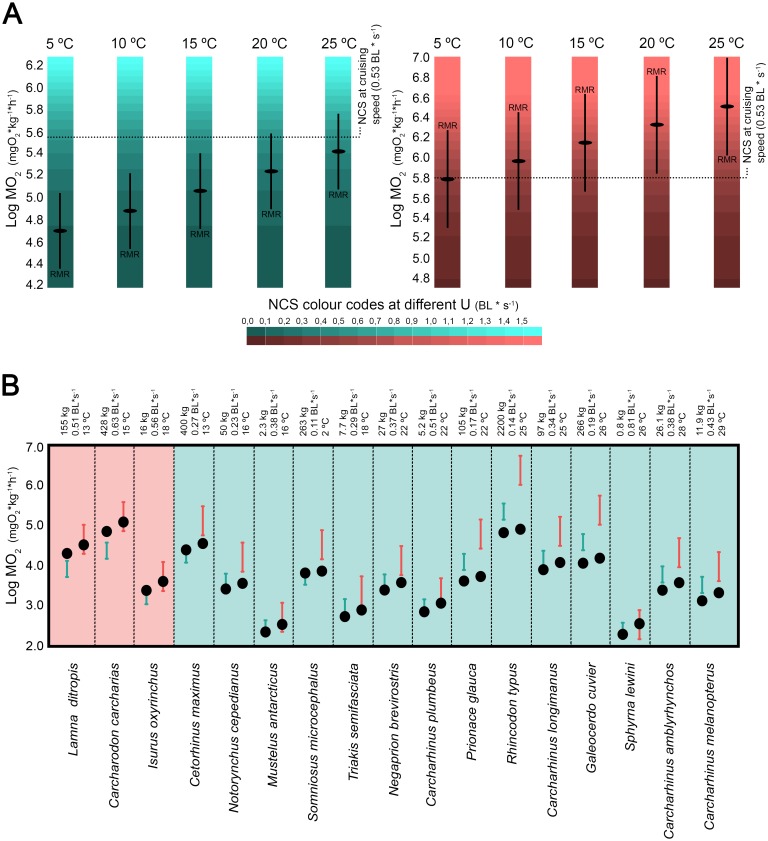
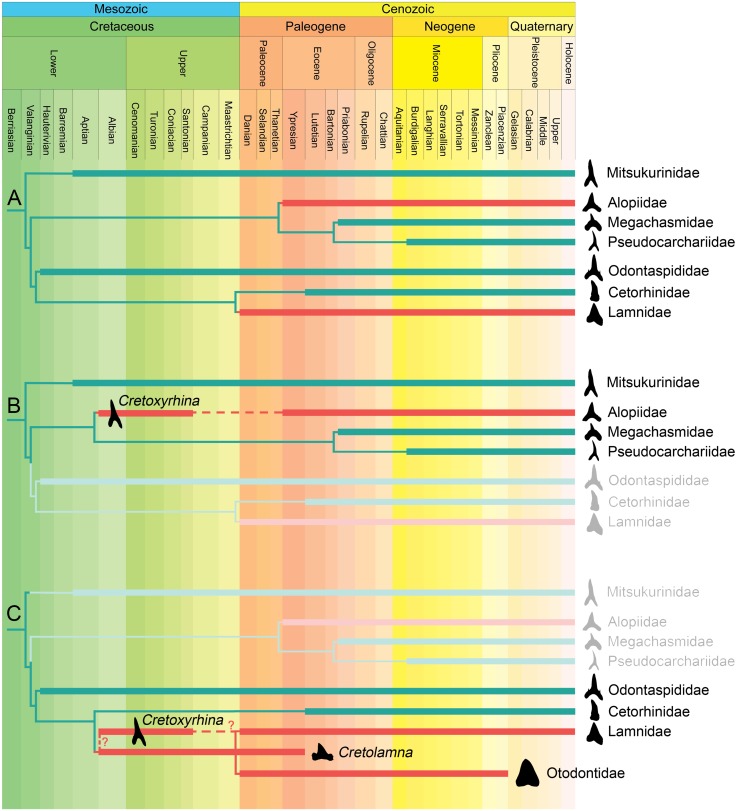
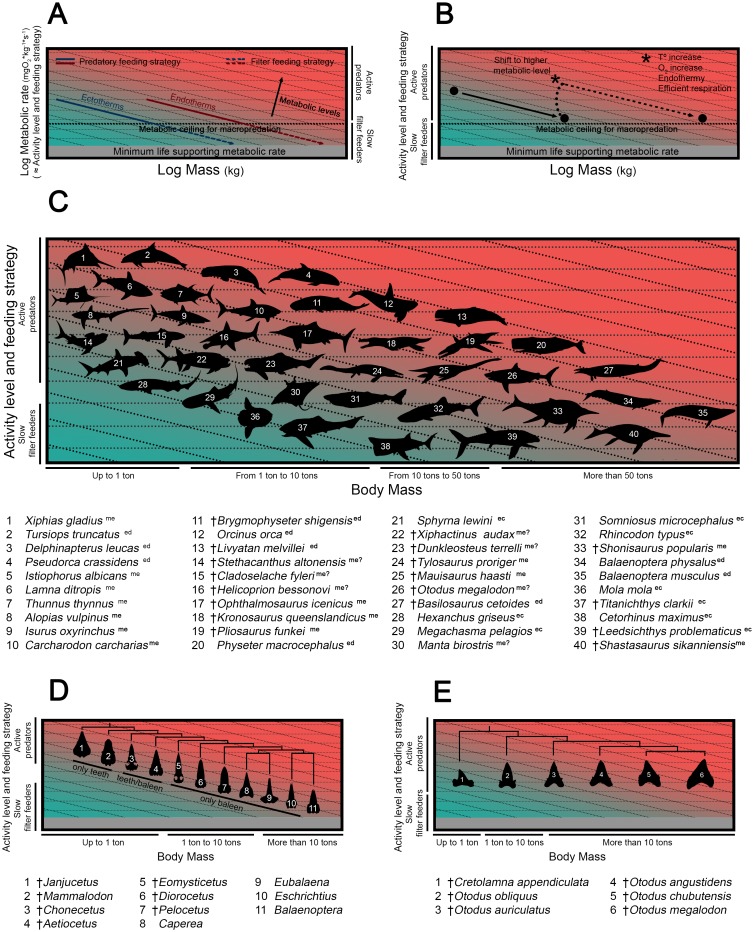
Otodontids include some of the largest macropredatory sharks that ever lived, the most extreme case being Otodus (Megaselachus) megalodon. The reasons underlying their gigantism, distribution patterns and extinction have been classically linked with climatic factors and the evolution, radiation and migrations of cetaceans during the Paleogene. However, most of these previous proposals are based on the idea of otodontids as ectothermic sharks regardless of the ecological, energetic and body size constraints that this implies. Interestingly, a few recent studies have suggested the possible existence of endothermy in these sharks thus opening the door to a series of new interpretations. Accordingly, this work proposes that regional endothermy was present in otodontids and some closely related taxa (cretoxyrhinids), playing an important role in the evolution of gigantism and in allowing an active mode of live. The existence of regional endothermy in these groups is supported here by three different approaches including isotopic-based approximations, swimming speed inferences and the application of a novel methodology for assessing energetic budget and cost of swimming in extinct taxa. In addition, this finding has wider implications. It calls into question some previous paleotemperature estimates based partially on these taxa, suggests that the existing hypothesis about the evolution of regional endothermy in fishes requires modification, and provides key evidence for understanding the evolution of gigantism in active macropredators.
Conflict of interest statement
Figures
Similar articles
-
Centralized red muscle in Odontaspis ferox and the prevalence of regional endothermy in sharks.Biol Lett. 2023 Nov;19(11):20230331. doi: 10.1098/rsbl.2023.0331. Epub 2023 Nov 8. Biol Lett. 2023. PMID: 37935371 Free PMC article.
-
Endothermic physiology of extinct megatooth sharks.Proc Natl Acad Sci U S A. 2023 Jul 4;120(27):e2218153120. doi: 10.1073/pnas.2218153120. Epub 2023 Jun 26. Proc Natl Acad Sci U S A. 2023. PMID: 37364100 Free PMC article.
-
Evolutionary pathways toward gigantism in sharks and rays.Evolution. 2019 Mar;73(3):588-599. doi: 10.1111/evo.13680. Epub 2019 Jan 24. Evolution. 2019. PMID: 30675721
-
Review: Analysis of the evolutionary convergence for high performance swimming in lamnid sharks and tunas.Comp Biochem Physiol A Mol Integr Physiol. 2001 Jun;129(2-3):695-726. doi: 10.1016/s1095-6433(01)00333-6. Comp Biochem Physiol A Mol Integr Physiol. 2001. PMID: 11423338 Review.
-
Evolution and consequences of endothermy in fishes.Physiol Biochem Zool. 2004 Nov-Dec;77(6):998-1018. doi: 10.1086/423743. Physiol Biochem Zool. 2004. PMID: 15674772 Review.
Cited by
-
Centralized red muscle in Odontaspis ferox and the prevalence of regional endothermy in sharks.Biol Lett. 2023 Nov;19(11):20230331. doi: 10.1098/rsbl.2023.0331. Epub 2023 Nov 8. Biol Lett. 2023. PMID: 37935371 Free PMC article.
-
Dental geochemistry reveals thermoregulation in the Neogene ocean's most infamous superpredator.Proc Natl Acad Sci U S A. 2023 Jul 25;120(30):e2308015120. doi: 10.1073/pnas.2308015120. Epub 2023 Jul 13. Proc Natl Acad Sci U S A. 2023. PMID: 37440567 Free PMC article. No abstract available.
-
White shark comparison reveals a slender body for the extinct megatooth shark, Otodus megalodon (Lamniformes: Otodontidae).Palaeontol Electronica. 2024 Jan 1;27(1):a7. doi: 10.26879/1345. eCollection 2024 Jan 1. Palaeontol Electronica. 2024. PMID: 39404696 Free PMC article.
-
Cautionary tales on the use of proxies to estimate body size and form of extinct animals.Ecol Evol. 2024 Sep 2;14(9):e70218. doi: 10.1002/ece3.70218. eCollection 2024 Sep. Ecol Evol. 2024. PMID: 39224151 Free PMC article.
-
The Early Pliocene extinction of the mega-toothed shark Otodus megalodon: a view from the eastern North Pacific.PeerJ. 2019 Feb 13;7:e6088. doi: 10.7717/peerj.6088. eCollection 2019. PeerJ. 2019. PMID: 30783558 Free PMC article.
References
-
- Capetta H. Chondrichthyes. Mesozoic and Cenozoic Elasmobranchii: Teeth Handbook of Paleoichthyology Volume 3E Verlag Dr Friedrich Pfeil, München; 2012.
-
- Leriche M. Les poissons Neogenes de la Belgique. Mémoires du Musée royal d’histoire naturelle de Belgique. 1926;32: 365–472.
-
- Bendix-Almgreen SE. Carcharodon megalodon from the Upper Miocene of Denmark, with comments on elasmobranch tooth enameloid: coronoïn. Bulletin of the Geological Society of Denmark. 1983;32: 1–32.
-
- Uyeno T, Sakamoto O, Sekine H. The description of an almost complete tooth set of Carcharodon megalodon from a Middle Miocene bed in Saitama Prefecture, Japan. Bulletin—Saitama Museum of Natural History. 1989;7: 73–85.
-
- Ehret DJ. Paleobiology and taxonomy of extinct lamnid and otodontid sharks (Chondrichthyes, Elasmobranchii, Lamniformes). Ph.D. thesis, University of Florida. 2010.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
