

A Spatial Interactome Reveals the Protein Organization of the Algal CO2-Concentrating Mechanism
- PMID: 28938113
- PMCID: PMC5616186
- DOI: 10.1016/j.cell.2017.08.044
A Spatial Interactome Reveals the Protein Organization of the Algal CO2-Concentrating Mechanism
Abstract
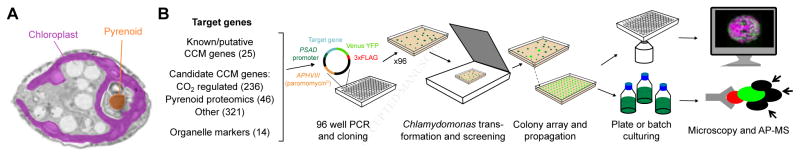
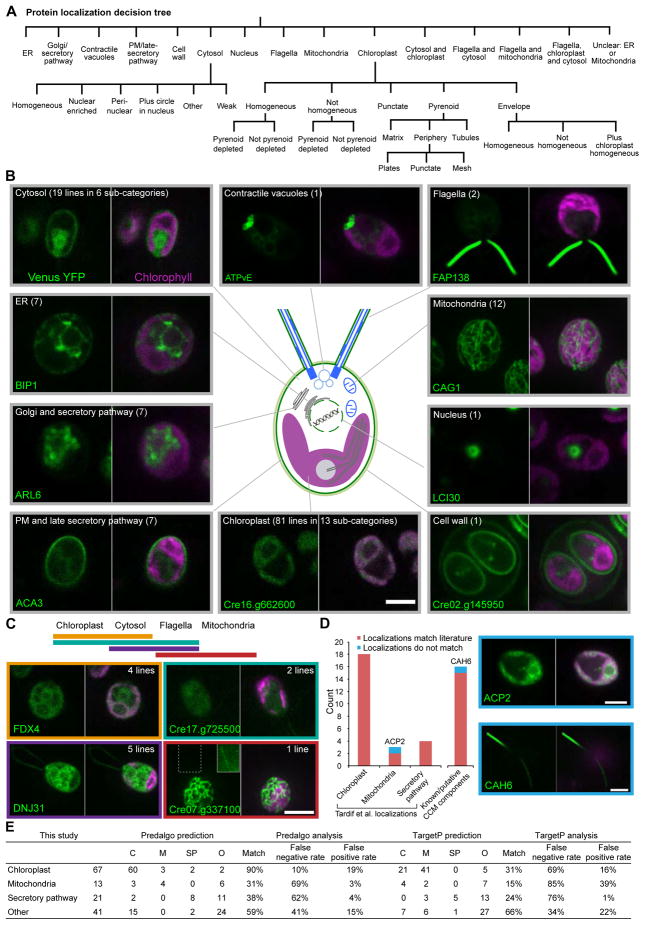
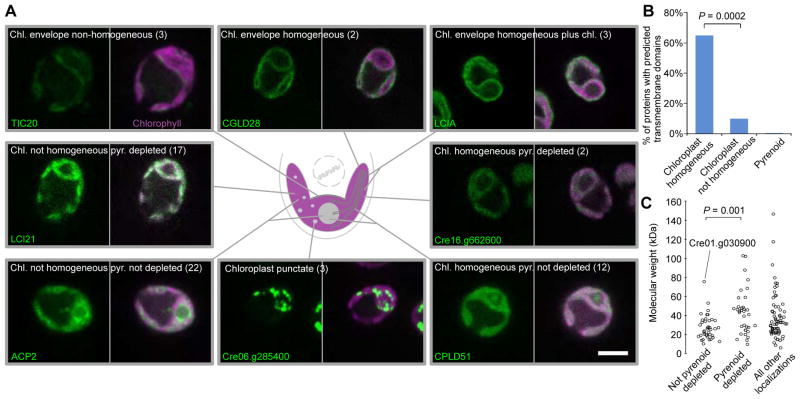
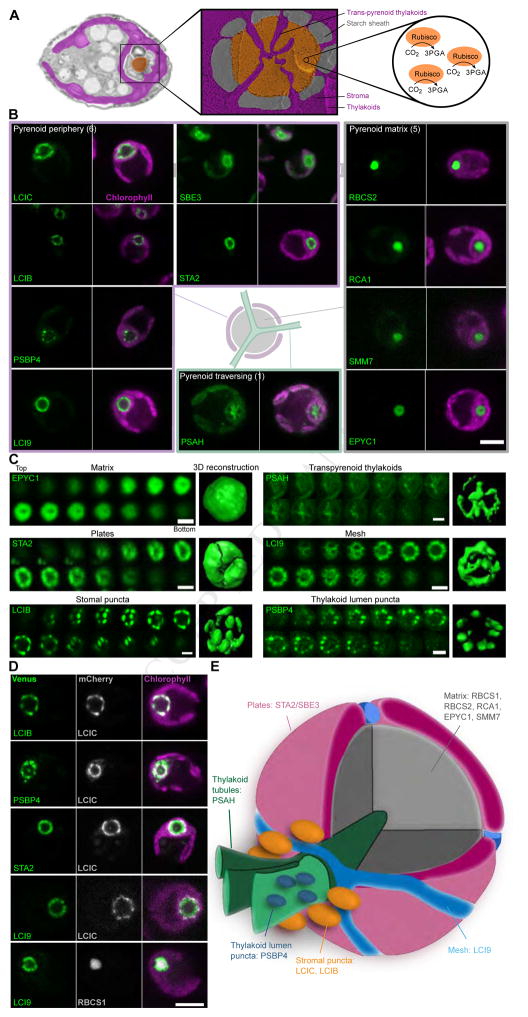
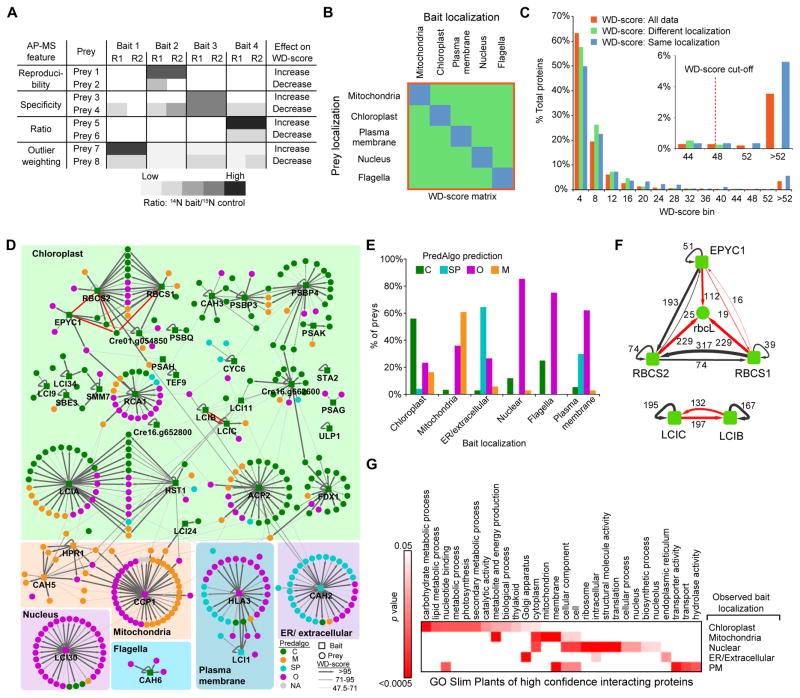
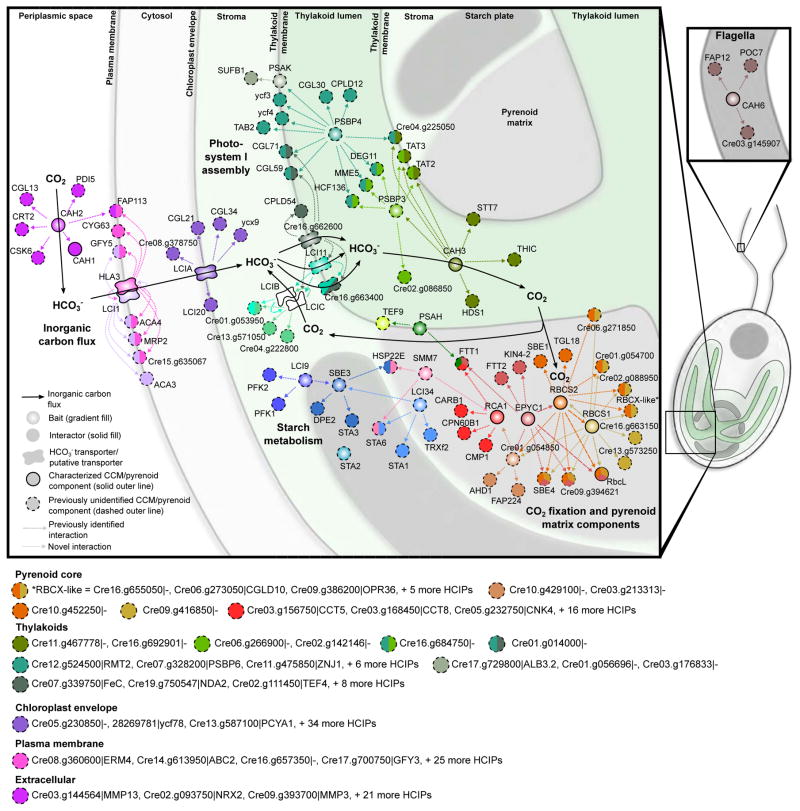
Approximately one-third of global CO2 fixation is performed by eukaryotic algae. Nearly all algae enhance their carbon assimilation by operating a CO2-concentrating mechanism (CCM) built around an organelle called the pyrenoid, whose protein composition is largely unknown. Here, we developed tools in the model alga Chlamydomonas reinhardtii to determine the localizations of 135 candidate CCM proteins and physical interactors of 38 of these proteins. Our data reveal the identity of 89 pyrenoid proteins, including Rubisco-interacting proteins, photosystem I assembly factor candidates, and inorganic carbon flux components. We identify three previously undescribed protein layers of the pyrenoid: a plate-like layer, a mesh layer, and a punctate layer. We find that the carbonic anhydrase CAH6 is in the flagella, not in the stroma that surrounds the pyrenoid as in current models. These results provide an overview of proteins operating in the eukaryotic algal CCM, a key process that drives global carbon fixation.
Keywords: CCM; CO(2)-concentrating mechanism; Chlamydomonas reinhardtii; Rubisco; affinity purification mass spectrometry; carbon fixation; high-throughput fluorescence protein tagging; photosynthesis; pyrenoid.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures

Comment in
-
The Pyrenoid: An Overlooked Organelle Comes out of Age.Cell. 2017 Sep 21;171(1):28-29. doi: 10.1016/j.cell.2017.09.012. Cell. 2017. PMID: 28938119
References
-
- Atteia A, Adrait A, Brugière S, Tardif M, Van Lis R, Deusch O, Dagan T, Kuhn L, Gontero B, Martin W. A proteomic survey of Chlamydomonas reinhardtii mitochondria sheds new light on the metabolic plasticity of the organelle and on the nature of the α-proteobacterial mitochondrial ancestor. Mol Biol Evol. 2009;26:1533–1548. - PubMed
-
- Badger M. The roles of carbonic anhydrases in photosynthetic CO2 concentrating mechanisms. Photosynthesis Res. 2003;77:83–94. - PubMed
-
- Bauwe H, Hagemann M, Fernie AR. Photorespiration: players, partners and origin. Trends Plant Sci. 2010;15:330–336. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
